
Dalbergia antsirananae Phillipson, Crameri & N. Wilding, 2021
|
publication ID |
https://doi.org/ 10.15553/c2021v762a4 |
|
persistent identifier |
https://treatment.plazi.org/id/03CBE223-224A-DB3C-3F0F-FAB6FA02A93D |
|
treatment provided by |
Felipe |
|
scientific name |
Dalbergia antsirananae Phillipson, Crameri & N. Wilding |
| status |
sp. nov. |
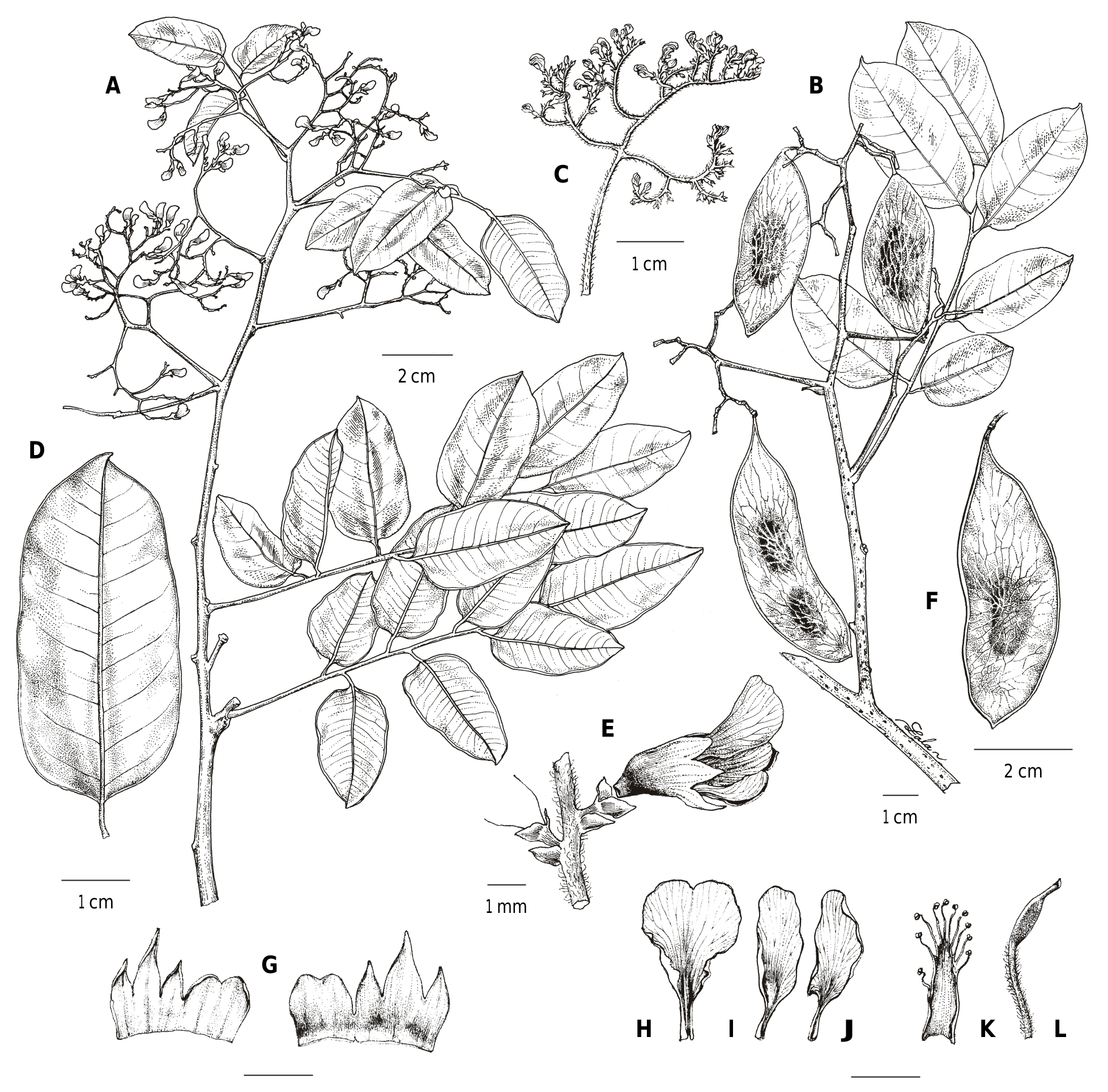
Dalbergia antsirananae Phillipson, Crameri & N. Wilding , sp. nov. ( Fig. 2 View Fig , 3 View Fig ).
Holot y pus: MADAGASC AR. Reg. DIANA [Pro v. Antsiranana]: Montagne des Français , 30.III.1994, fl., Du Puy & Andriantiana M761 ( P [ P00059481 ]!; iso-: K [ K001351089 ]!, MO-3065175 image!, PRE, TAN, WAG [ WAG1014216 View Materials , WAG1014217 View Materials 2-sheet specimen] images!) .
Dalbergia antsirananae Phillipson, Crameri & N. Wilding differs from D. madagascariensis Vatke by its smaller flowers (petal length c. 5 mm vs. at least 6 mm) and leaves with (5–)7– 9 membranous, ovate-triangular leaflets, each with a shortly acuminate apex (vs. 5–7(–9) subcoriaceous, ovateoblong leaflets, each with an attenuate apex).
Trees to c. 15 m tall; bole to 3.5 m high, DBH to c. 50 cm. Branches glabrous, yellow-green in vivo (black in sicco) when young, becoming pale grey-brown when old, coppice shoots with a fine brown pubescence on the branches and the leaves (RIR, pers. obs.; Fig. 2C View Fig ), lenticels present. Leaves alternate,
[A: Randrianaivo 3675; B: Razafitsalama1504; C–D: Randrianaivo 3506] [Photos: A, C–D: R. Randrianaivo; B: J. Razafitsalama]
c. 12–20 cm long, with (5–)7–9 alternate leaflets; petiole and rachis yellow-green in vivo, dark brown to black in sicco, glabrous except for a very fine, scattered, white, puberulous indument forming a line on the upper surface of the rachis, with a fine brown pubescence on coppice shoots; petiole 2.5–3.5 cm long; stipules caducous (none seen on available specimens), leaving a visible scar; leaflets (5–)40–55(–70) × (3–)20–25(–30) mm, the proximal leaflets sometimes smaller than the rest, the distal leaflet slightly larger, the others often relatively uniform; petiolule (2–) 3.5–5 mm long, yellow-green in vivo, very dark brown to black in sicco, with a very fine, scattered, white, puberulous indument on the upper surface, with a fine brown pubescence on the lower surface on coppice shoots; lamina ovate-triangular, membranous, base truncate to broadly obtuse, margin very slightly revolute, apex acute to obtuse, shortly acuminate, venation brochidodromous, with 7–9 principal lateral veins per side; upper and lower surface bright mid-green in vivo, dark brown in sicco, upper surface slightly glossy, glabrous, the midrib forming a groove, with a very fine, scattered, white, puberulous indument on the proximal portion, lower surface matt, glabrous, higher-order veins forming a fine network, midrib prominent. Inflorescences terminal, corymbose panicles, corymbs
2 mm
2 mm
adaxial surface); L. Gynoecium.
[A, C–E, G–L: Service Forestier 13215, TEF; B: Randriambololomamonjy 23, TAN; F: Randrianarivelo et al. 138, TAN] [Drawings:R.L. Andriamiarisoa]
decreasing in size towards the apex, the basal units subtended by well-developed pinnate leaf-like bracts, the apical units with reduced, caducous scale-like bracts or caducous leafy bracts, distal portions curved, sub-spicate, secundiflorous, axes sparsely pubescent; peduncle 2–3 cm long. Flowers subtended by a caducous, triangular, sparsely puberulous (mostly near the margins) bract, c. 0.5–1.5 × 0.5–1.5 mm; pedicel 0.5–1.2 mm long, very sparsely puberulous or glabrous (pedicels of apical flowers shorter than of basal flowers); bracteoles ovate, c. 1.2 × 1 mm, apex rounded, sparsely ciliate, yellow-green in vivo, brown to dark brown in sicco, caducous; calyx yellow-green in vivo (or cream fide Du Puy M761), becoming brown to dark brown in sicco, glabrous except for minutely ciliate apices of lower 3 lobes, persisting on fruits, c. 3 mm long, fused in the lower 2/3, the 2 upper lobes fused to form a single, rounded unit, c. 2 × 3 mm, the apex shallowly emarginate, the 2 lateral lobes triangular, c. 1 × 1 mm, the lowest lobe triangular, keeled, c. 1.8 × 1.4 mm; petals glabrous, white in vivo, becoming pale brown in sicco; standard petal pandurate, c. 5 × 3 mm, claw c. 1 mm long, apex notched; wing petals c. 5 × 1.2 mm, claw c. 0.8 mm long; keel petals c. 5 × 1.5 mm, claw c. 1.8 mm long; androecium glabrous, monadelphous, c. 4 mm long; stamens 10, filaments free for the upper 1/3; gynoecium c. 4.7 mm long; stipe densely pubescent, c. 1.9 mm long; ovary glabrous, c. 2 mm long, with 4 ovules; style glabrous, c. 0.8 mm long. Fruits green in vivo (fide Rakotondrajaona 316), dark brown in sicco, the body oval, 5.0–6.0 × 1.9–2.1 cm when single-seeded (multi-seeded fruits not seen), base cuneate, apex rounded, surface smooth but with a coarse network of mostly longitudinal ribs over the seed not extending to the edges, glabrous; stipe c. 4 mm long, glabrescent but with some persistent indument at the base; style caducous; seeds 7 × 3.5 mm, reniform, flattened, brown, smooth.
Etymology. – The name Dalbergia antsirananae was chosen to reflect the species’ restricted distribution near the city of Antsiranana in northern Madagascar.
Vernacular names and uses. – “Ma(g)nary” (Du Puy M761, Randriamahazomanana 130, Service Forestier 13215); “Tsiandalana” (Randriambolomamonjy 12, Randrianarivelo et al. 138).
The wood of Dalbergia antsirananae is strong and durable, and red-brown in colour; it is used in carpentry and for furniture, and it is also reported as good for charcoal production (Du Puy & Andriantiana M761).
Distribution, ecology and phenology. – Dalbergia antsirananae occurs in intact and degraded, dry, dense deciduous forest and gallery forest at low- to mid-elevations (55– 450 m) in the north of Madagascar. It occurs mainly on tertiary limestone (tsingy) and on alluvial or white sandy soils over limestone, but also on basaltic substrates. The species has been collected in flower from April to May, just after the rainy season, and in fruit in June and from September to October, during the dry season. A single record of young fruit in March 2004 at the start of the dry season is probably due to exceptionally dry weather in February of that year ( SZABÓ et al., 2015).
Conservation status. – Dalbergia antsirananae has an EOO of c. 128 km ² and an estimated minimum AOO of 36 km ² (based on a 4 km ² grid). The species occurs in two protected areas: the Montagne des Français (Ambohitratsingy) Protected Landscape, a category V protected area, where it has been documented by nine collections at scattered localities, and by seven collections from two separate low-lying areas in the north of the Montagne d’Ambre National Park, a category II protected area, that are atypical of the National Park as a whole, most of which is over 500 m in elevation and covered primarily by humid forest and with little, if any, dry forest. These three areas represent three separate subpopulations. Within the Montagne des Français protected area, where the largest sub-population of D. antsirananae occurs, the species is threatened by wild fires related to charcoal production, as exemplified by a fire that destroyed a 4 ha block of forest in November 2020 within the protected area (C. Frasier, pers. comm.), while illegal timber harvesting may also occur. Recent observations of the two subpopulations in Montagne d’Ambre National Park have provided valuable information on their status. The species is abundant at one of the sites (RIR, pers. obs., Feb. 2021), and this important subpopulation should be targeted by park authorities for conservation measures because it is easily accessed and therefore particularly vulnerable. The second subpopulation within the National Park is less accessible and the species apparently less abundant (RIR, pers. obs., Oct. 2019), but at this site a large, recently felled tree of D. antsirananae with a bole of c. 50 cm diameter was documented in October 2019 (Randrianaivo 3506; Fig. 2D View Fig ) although coppice shoots were developing from its base. With respect to wild fires, which are considered to be the most serious plausible threat, D. antsirananae occurs at three locations corresponding to the three known subpopulations, and on this basis it is assigned a preliminarily conservation status of “Endangered” [EN B1ab(i,ii,iii,iv,v)+2ab(i,ii,iii,iv,v)] following the IUCN Red List Categories and Criteria (IUCN, 2012). The impact of recent fires at the Montagne des Français on this important subpopulation should be evaluated, and the three subpopulations should be closely monitored.
Notes. – The currently available material of Dalbergia antsirananae includes two collections that were seen by Bosser and Rabevohitra in 1992 (Service Forestier 3557, 13215) and one (Du Puy & Andriantiana M761) examined by Bosser in 1997. These specimens were identified and annotated by Bosser as D. madagascariensis , and no doubt for this reason, in their treatment of the genus, BOSSER & RABEVOHITRA (2002) indicated that D. madagascariensis occurred as far north as the city of Antsiranana. However, these and other specimens now available from the same area are morphologically distinct from D. madagascariensis (see Table 1), which, as now circumscribed, does not occur in this area.
Differences in habitat further support the recognition of Dalbergia antsirananae as a coherent taxonomic entity distinct from D. madagascariensis : D. antsirananae occurs in dry deciduous forest whereas D. madagascariensis occurs is humid and sub-humid evergreen forest. In addition to differences in the morphology of the flowers and leaves of D. antsirananae , which separate it from D. madagascariensis (see diagnosis), it can further be distinguished from its allies by a combination of vegetative and floral characters (see Table 1).
Additional specimens examined. – MADAGASCAR. Reg. DIANA [Prov. Antsiranana]: Distr. d’Antsiranana II, Montagne des Français PA ( Paysage Harmonieux Protégé d’Ambohitr’Antsingy), 30.III.2004, y.fr., Rakotondrajaona et al. 316 ( CNARP, MO, P, TAN) ; ibid. loco, 14.XII.2019, ster., Ramanitrinizaka & Ravaomanalina 158 ( DBEV, MO, P, TAN, ZT) ; ibid. loco, 12. VI.2019, fr., Randriamahazomanana 130 ( MO, P, TAN) ; ibid. loco, 5.X.2004, fr., Randriambololomamonjy 23 ( MO, P, TAN) ; ibid. loco, 24. I.2014, ster., Randrianaivo et al. 2451 ( BR, G, MO, P) ; ibid. loco, 25.II.2021, fl., Randrianaivo & Andriamiadana 3675 ( DBEV, MO, P, TAN, ZT) ; ibid. loco, fl., Randrianaivo & Andriamiadana 3676 ( DBEV, MO, P, TAN, ZT) ; ibid. loco, 24.IX.2004, fr., Randrianarivelo et al. 138 ( MO, P, TAN) ; Montagne d’Ambre NP, Antsampano , 31.X.2019, ster., Randrianaivo & Andriamiadana 3506 ( DBEV, MO, P, TAN, ZT) ; Montagne d’Ambre NP, Sakaramy , 28.II.2021, fl., Randrianaivo & Andriamiadana 3682 ( DBEV, MO, P, TAN, ZT) ; ibid. loco, fl., Randrianaivo & Andriamiadana 3683 ( DBEV, MO, P, TAN, ZT) ; ibid.loco, fl., Randrianaivo & Andriamiadana 3684 ( DBEV, MO, P, TAN, ZT) ; ibid. loco, fl., Randrianaivo & Andriamiadana 3685 ( DBEV, MO, P, TAN, ZT) ; ibid. loco, 29.IV.2021, fr., Razafitsalama 1504 ( MO, P, TAN) ; Montagne des Français , 21. V.1951, fl., Service Forestier 3557 ( P) ; Montagne d’Ambre, Sakaramy , 3.IV.1955, fl., Service Forestier 13215 ( G, K, MO, P, TEF, WAG) .
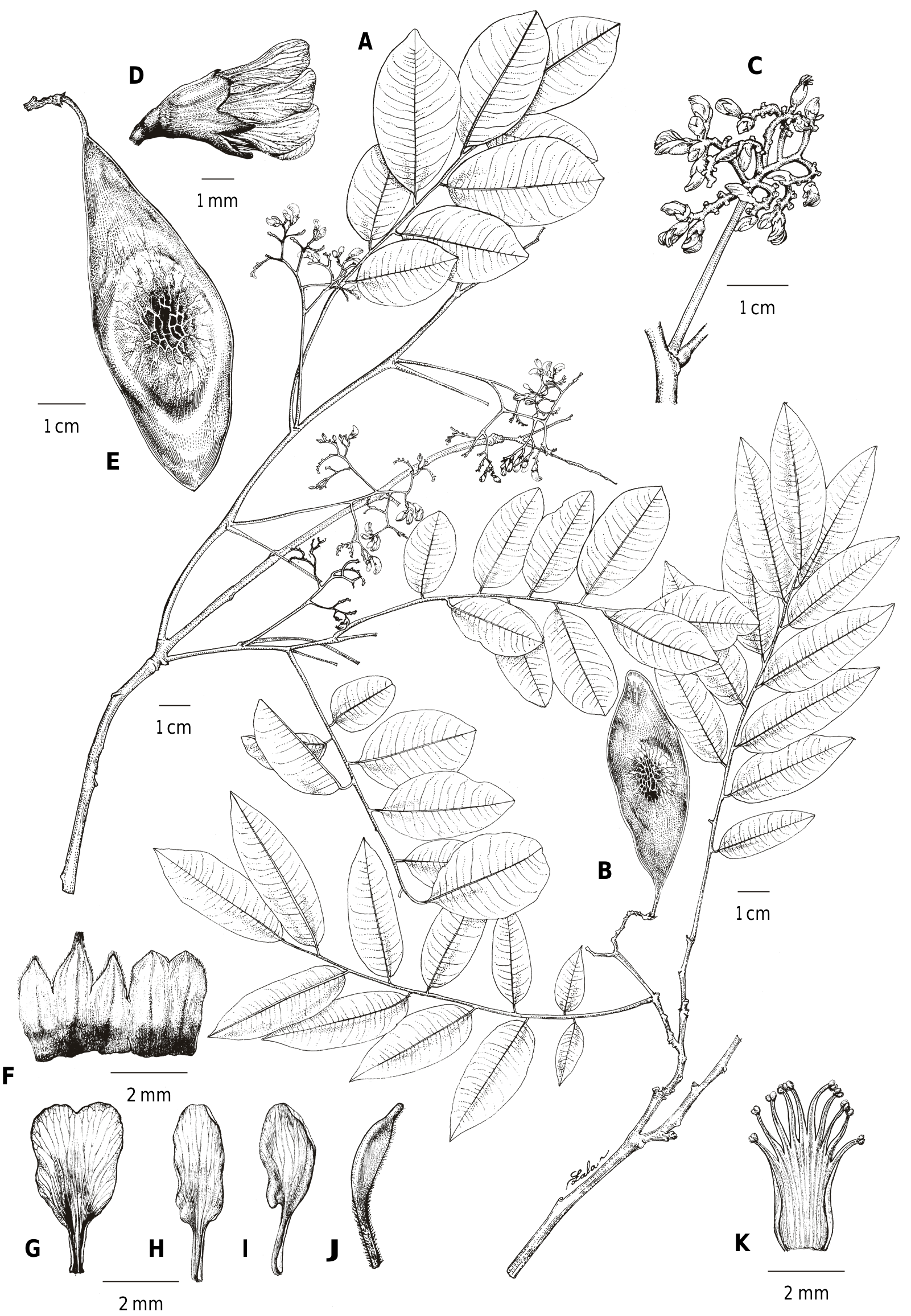
Dalbergia manongarivensis Bosser & R. Rabev. View in CoL in Adansonia ser. 3, 27: 210. 2005 ( Fig. 4 View Fig ).
H o l o t y p u s: M A D A G A S C A R. R e g. S o f i a [ P r o v. Mahajanga]: Manongarivo SR, 3.VI.1996, fl., Totozafy Be 537 ( G [ G00011766 ] image!; iso-: K [ K001351163 ]!, MO-4924628!, P [ P00513688 ]!, TEF) .
Trees or shrubs (or possibly lianescent shrubs) to c. 22 m tall; bole to 8 m high, DBH to c. 25 cm. Branches glabrous, green to olive-green in vivo (brown in sicco) when young, becoming pale grey-brown when old, lenticels present. Leaves alternate, c. 16–20.5 cm long, with 7–11(–12) alternate leaflets; petiole and rachis pale green in vivo, dark brown in sicco, glabrous except for a fine, scattered, white, puberulous indument on the entire surface of the rachis; petiole 2.5–4.5 cm long; stipules caducous (none seen on available specimens), leaving a visible scar; leaflets (1.5–)4–8.5 × (1–) 1.5–3 cm, the proximal 2 or 3 leaflets often smaller than the rest, the distal 2 or 3 often larger; petiolule 2–3 mm long, pale green in vivo, dark brown in sicco, with a fine, scattered, white, puberulous indument on the upper surface; lamina elliptic, membranous, base rounded to obtuse, margin slightly revolute, apex acute to obtuse, shortly acuminate, venation brochidodromous, with c. 10 principal lateral veins per side; upper and lower surface bright mid-green in vivo, mid to dark brown in sicco, upper and related species occurring in northern Madagascar.
D. antsirananae D. madagascariensis D. manongarivensis D. viguieri number (5–)7–9 5–7(–9) 7–11(–12) 5–7
lamina shape ovate-triangular ovate-oblong elliptic elliptic-suborbicular Leaflets lamina texture membranous subcoriaceous membranous membranous acute to obtuse, acute to obtuse, apex acute to
lamina apex attenuate
shortly acuminate shortly acuminate rounded position terminal terminal or axillary axillary axillary
Inflorescences length
as long as or longer as long as or longer
shorter than leaves shorter than leaves
than leaves than leaves length [mm] 5 6–7.5(–9) 5 (3.5–)4–5
Flowers calyx length [mm] 3 3–5(–6.5) 4 2.5–3
elliptic-oblong, elliptic-oblong,
elliptic, 3–4 × 1–1.3
5–10 × 1.5–3 7.5–9 × 3.3
shape [cm] oval, 5–6 × 1.9–2.1 or 5–6 × 1–1.3
Fruits or 13–15 × 1.5–3 or c. 11.5 × 2.8 (2-seeded fruits) (2-seeded fruits) (2-seeded fruits)
stipe length [mm] c. 4 c. 4 c. 8 5–8
surface slightly glossy, glabrous, the midrib forming a groove, with a fine, scattered, white, puberulous indument on the proximal portion, lower surface matt, with scattered appressed indument, higher-order veins forming a fine network, midrib prominent, with indument present along sides. Inflorescences axillary, a series of corymbs, sometimes appearing terminal due to damaged or aborted terminal growth, the successive corymbs of similar size (not noticeably decreasing towards the apex of the fertile branch), subtended by a well-developed pinnate leaf-like bract and with reduced, caducous, scale-like bracts present within the corymb, distal portions curved, subspicate, secundiflorous, axes glabrous to sparsely pubescent; peduncle c. 4–8 cm long. Flowers subtended by a caducous, triangular, ciliate bract, c. 0.5–2 × 0.5–2 mm; pedicel c. 1 mm long, glabrous to sparsely pubescent; bracteoles ovate, c. 1.5 × 1.5 mm, apex rounded, sparsely ciliate, brown in sicco, caducous; calyx pale green in vivo (Nusbaumer 2564), becoming dark brown in sicco, villous or glabrous except for sparsely and minutely ciliate margins, persisting on fruits, c. 4 mm long, fused in the lower half, the 2 upper lobes fused to form a single, rounded unit, c. 1.5 × 2 mm, apex shortly notched, the 2 lateral lobes rounded-triangular, c. 1.5 × 1 mm, the lowest lobe rounded-triangular, keeled, c. 2.2 × 1 mm, with a distinctly curved, apex attenuate; petals glabrous, white in vivo, becoming yellow-brown in sicco; standard petal pandurate, c. 5 × 3 mm, claw c. 1 mm long, apex notched; wing petals c. 5 × 1.4 mm, claw c. 1 mm long; keel petals c. 5 × 1.3 mm, claw c. 1.8 mm long; androecium glabrous, monadelphous, c. 4.5 mm long; stamens 10, filaments free for the upper 2/5; gynoecium c. 4.8 mm long; stipe densely pubescent, c. 2.1 mm long; ovary glabrous, c. 1.8 mm long, with 4 ovules; style glabrous, c. 0.9 mm long. Fruits pale yellow-green in vivo (Karatra 159), dark brown in sicco, remnants of the petals and calyx sometimes persisting at base of fruits, the body narrowly elliptic-rhombic, c. 7.5– 9 × 3.3 cm, when single-seeded, or narrowly elliptic-oblong, c. 11.5 × 2.8 cm, when multi-seeded, base acute, apex acute, surface smooth but with coarse network of fine, mostly longitudinal ribs over the seed, and extending to the edges, glabrous; stipe c. 8 mm long, glabrescent but sometimes with indument persisting at the base; style caducous; seeds (immature) 9 × 5 mm, sub-reniform, flattened, brown, smooth.
Vernacular names and uses. – “ Faralabikesa” (Ammann et al. 391, Tahinarivony et al. 708); “Hazovola” (Randriatsivery 446, Reserves Naturelles 6911, Service Forestier 7710, Wohlhauser 60279); “Manary” (Perrier 15188); “Manary Bomby” (“Manaibomby”) (Antilahimena 404, 805); “Manary Boraka” (Service Forestier 3100, 95R152 [Tsimihety dialect]).
Specific uses do not appear to have been recorded for any of the available herbarium specimens of Dalbergia manongarivensis , however wood apparently from trees of this species from Manongarivo, wrongly determined and illustrated as D. madagascariensis , has been described as a valued source of timber ( ANONYMOUS, 2012: 40–44, photo Nº 12).
Distribution, ecology and phenology. – Dalbergia manongarivensis occurs in primary lowland moist evergreen forest and disturbed areas in the Sambirano biogeographic region of northwestern Madagascar, from near sea level to c. 700 m, on “rocky ground”, alluvial soils, and gneiss. It is frequently reported as occurring on the edge of rivers and streams and on “lower slopes”. Some of specimen labels for collections from Ankaramy indicate DIANA Region or Antsiranana Province, but the locality details and/or coordinates clearly indicate that these collections were in fact made within the Sofia Region (Mahajanga Province); the data have been corrected accordingly. Dalbergia manongarivensis has been collected in flower from March to July and from October to December, and in fruit from July to November and in January. This corresponds most closely to periods of drier and slightly cooler conditions within its known distribution range; precipitation is typically at its lowest from April to October.
Conservation status. – Dalbergia manongarivensis has an estimated EOO of c. 3900 km ² and a minimum AOO of 104 km ² (based on a 4 km ² grid). The species occurs within four formally recognised protected areas: Manongarivo Special Reserve (category II), Tsaratanana Strict Nature Reserve (category I), and both the Ampasindava and Galoko-Kalobinono Sustainable Use Areas (category V), as well as in unprotected areas between these sites. In the latter, as well as in less well-managed protected areas, the species is subject to land clearance for agriculture resulting in a loss of suitable habitat, which we consider to be the most serious plausible threat. On this basis, D. manongarivensis occurs at 13 locations, and is assigned a preliminary conservation status of “Near Threatened” [NT] according to Criterion B of the IUCN Red List criteria (IUCN, 2012). In unprotected areas, as well as at Ampasindava and Galoko-Kalobinono, exploitation of timber resources is permitted and consequently D. manongarivensis is also subjected to logging. Based on the Criterion A approach outlined above, we suspect a population size reduction of 77 % for D. manongarivensis over the period 1990–2089, based on IUCN Criterion A4(c), i.e. “a decline in habitat quality”, using a generation time of 33 years, and assuming no habitat loss within protected areas (IUCN Categories I–IV). Dalbergia manongarivensis is therefore assigned a preliminary conservation status of “Endangered” [EN A4(c)] following the IUCN Red List Categories and Criteria (IUCN, 2012). The assessment under Criterion A suggests a higher level of threat of extinction than the assessment under Criterion B and therefore it should be used in a formal assessment for this species.
F. Calyx (outer surface; split open and flattened); G. Standard petal (adaxial surface); H. Wing petal (adaxial surface); I. Keel petal (adaxial surface); J. Gynoecium; K. Androecium (split open to show adaxial surface).
[A,F–K: Antilahimena 404, TAN; B: Birkinshaw 532 TAN; C–D: Randriatsivery 437 (ex photo); E: Randriatsivery 446, TAN] [Drawings: R.L. Andriamiarisoa]
Notes. – Dalbergia manongarivensis was described by BOSSER & RABEVOHITRA (2005) in their final published taxonomic contribution on the genus. It was based on a single collection from Manongarivo (Totozafy Be 537), which was described on the corresponding herbarium label in 1996 as a woody liana (“liane ligneuse”). Despite extensive inventory effort at Manongarivo and in its surroundings, and also at Galoko and Ampasindava, which has generated a wealth of new Dalbergia collections, no additional lianescent material has ever been recorded. Numerous specimens of arborescent Dalbergia from Manongarivo were identified by Bosser & Rabevohitra as D. madagascariensis , a species for which they adopted a very broad circumscription (BOSSER & RABEVOHITRA, 2002) that also included some of the material now assigned to the newly described D. antsirananae (see above). Phylogenomic and population genomic results ( CRAMERI, 2020) support the recognition of arboreal forms (e.g. Ammann et al. 391, Antilahimena 404, Randriatsivery et al. 437, 446) and the single supposedly lianescent collection (Totozafy Be 537) as a coherent entity that is clearly differentiated from D. madagascariensis . This conclusion is supported by the morphological similarity of the leaves and inflorescences of arborescent specimens and those of the lianescent type collection. As circumscribed here, D. manongarivensis is clearly distinguished from related species (see Table 1). The observation of a lianescent habit on the label of Totozafy Be 537 was possibly an error made in the field or when transcribing the field notes, but it is also possible that both lianescent and arborescent or shrubby forms of D. manongarivensis do exist. According to some accounts (KLITGAARD & LAVIN, 2005; LACHENAUD & VAN DER MAESEN, 2016), the growth form of individuals of certain species of Dalbergia may be influenced by environmental conditions. This observation is based on species from continental Africa and Asia, and further study is required to determine whether this phenomenon occurs in Madagascar.
Additional specimens examined. – MADAGASCAR Reg. DIANA [Prov. Antsiranana]: Ampasindava PA, 6.XII.2008, bud, fl., Ammann et al. 147 ( G, K, MO, P, TEF) ; ibid. loco, 26.11.2009, fl., Ammann et al. 391 ( G, K, MO, P, TEF) ; Tsaratanana RNI, 24.III.2000, bud, fl., Antilahimena 404 ( K, MO, P, TAN) ; ibid. loco, 14.XI.2001, fr., Antilahimena et al. 805 ( P) ; ibid. loco, 8.X.1998, fr., Birkinshaw et al. 532 ( MO, P, TAN) ; Massif du Manongarivo, 14. V.1995, fl., Gautier 2653 ( G, MO, P, TAN) ; Ampasindava, 29.VII.2019, fr., Karatra et al. 157 ( MO, P, TAN) ; ibid. loco, 29.VII.2019, fr., Karatra et al. 158 ( DBEV, MO, P) ; ibid. loco, 29.VII.2019, fr., Karatra et al. 159 ( P) ; ibid. loco, 29.VII.2019, y.fr., Karatra et al. 161 ( DBEV, MO, P) ; ibid. loco, 29.VII.2019, st., Karatra et al. 162 ( P) ; ibid. loco, 30.VII.2019, fl., Karatra et al. 166 ( DBEV, MO, P) ; ibid. loco, 27.11.2007, fl., Nusbaumer 2564 ( G, K, MO, P, TAN, TEF, WAG) ; vallée du Sambirano, 1.IX.1908, fr., Perrier de la Bâthie 4107 ( P) ; ibid. loco, Perrier de la Bâthie 4115 ( P) ; Massif du Manongarivo, 1.IV.1909, fl., Perrier de la Bâthie 4137 ( P) ; vallée du Sambirano, 1.XII.1922, fr., Perrier de la Bâthie 15140 ( P) ; ibid. loco, XII.1922 – I.1923, fl., y.fr., Perrier de la Bâthie 15188 ( P) ; Ampasindava PA, 23.VII.2019, ster., Ramanitrinizaka et al. 94 ( DBEV, P) ; ibid.loco, 24.VII.2019, y.fr., Ramanitrinizaka et al. 99 ( DBEV, P, TAN) ; Galoko-Kalobinono PA, 23.III.2009, bud, fl., Briggs et al. 284 ( K, MO, P, TAN) ; ibid. loco, 7.X.2013, bud, fl., Randriatsivery et al. 437 ( G, MO, P, TAN) ; ibid. loco, y.fr., Randriatsivery et al. 446 ( G, MO, P, TAN) ; ibid. loco, 14.IV.2000, bud, fl., Ravelonarivo et al. 1176 ( MO, P, TAN) ; Massif du Manongarivo, 18.XII.2019, bud, fl., Razakamalala et al. 8586 ( DBEV, MO, P, TAN) ; Massif Marovato Ambanja, 19.XII.1954, fl., Réserves Naturelles 6911 ( K, MO, P, TEF) ; Galoko-Kalobinono PA, 26.X.1952, y.fr., Service Forestier 6115 ( K, P, TEF) ; ibid. loco, 2.X.1953, fl., Service Forestier 7710 ( K, P, TEF) ; Massif du Manongarivo, 10–16.XII.1954, fl., Service Forestier 11480 ( BR, G, K, MO, P, TEF, WAG) ; Ampasindava PA, 23. I.2009, bud, Tahinarivony et al. 252 ( G, K, MO, P) ; Massif du Manongarivo, 31. V.2000, fl., Wohlhauser 60279 ( G, K, MO, P, TAN) . Reg. Sofia [Prov. Mahajanga]: Ankaramy (district de Maromandia), 11. VI.1923, fl., Decary 2168 ( P) ; ibid. loco, 31.VIII.1950, fl., Service Forestier 3100 ( K, P, TAN, TEF) ; Massif de Maromiandra, 9–11.XI.1952, ster., Service Forestier 95-R-152 ( P, TEF) .

| P |
Museum National d' Histoire Naturelle, Paris (MNHN) - Vascular Plants |
| K |
Royal Botanic Gardens |
| PRE |
South African National Biodiversity Institute (SANBI) |
| TAN |
Parc de Tsimbazaza |
| WAG |
Wageningen University |
| PA |
Universidade Federal do Oeste do Pará |
| MO |
Missouri Botanical Garden |
| ZT |
Eidgenössische Technische Hochschule Zürich |
| VI |
Mykotektet, National Veterinary Institute |
| I |
"Alexandru Ioan Cuza" University |
| BR |
Embrapa Agrobiology Diazothrophic Microbial Culture Collection |
| G |
Conservatoire et Jardin botaniques de la Ville de Genève |
| V |
Royal British Columbia Museum - Herbarium |
| TEF |
Centre National de la Recherche Appliquée au Developement Rural |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Dalbergia antsirananae Phillipson, Crameri & N. Wilding
| Wilding, Nicholas, Phillipson, Peter B., Crameri, Simon, Andriambololonera, Sylvie, Andriamiarisoa, Roger L., Andrianarivelo, Sandratra A. F., Bernard, Roger, Rakotonirina, Nivo, Rakotovao, Charles, Randrianaivo, Richard I., Razakamalala, Richard & Lowry Ii, Porter P. 2021 |
Dalbergia manongarivensis
| Bosser & R. Rabev. 2005: 210 |