
Rufocumbre lucasi, Dolibaina & Mielke & Casagrande, 2017
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4365.2.5 |
|
publication LSID |
lsid:zoobank.org:pub:D9EA4014-FE5B-4F23-86F8-9E2853A312C2 |
|
DOI |
https://doi.org/10.5281/zenodo.6032063 |
|
persistent identifier |
https://treatment.plazi.org/id/ADF19B01-2D4F-4C1D-8E20-9AD0410159AF |
|
taxon LSID |
lsid:zoobank.org:act:ADF19B01-2D4F-4C1D-8E20-9AD0410159AF |
|
treatment provided by |
Plazi |
|
scientific name |
Rufocumbre lucasi |
| status |
sp. nov. |
Rufocumbre lucasi sp. nov.
urn:lsid:zoobank.org:act:ADF19B01-2D4F-4C1D-8E20-9AD0410159AF Figs 17–20 View FIGURES 1–20 , 25 View FIGURES 21–25 , 37 View FIGURES 35–37 , 42 View FIGURES 40–42 , 47 View FIGURES 43–47 , 56 View FIGURE 56
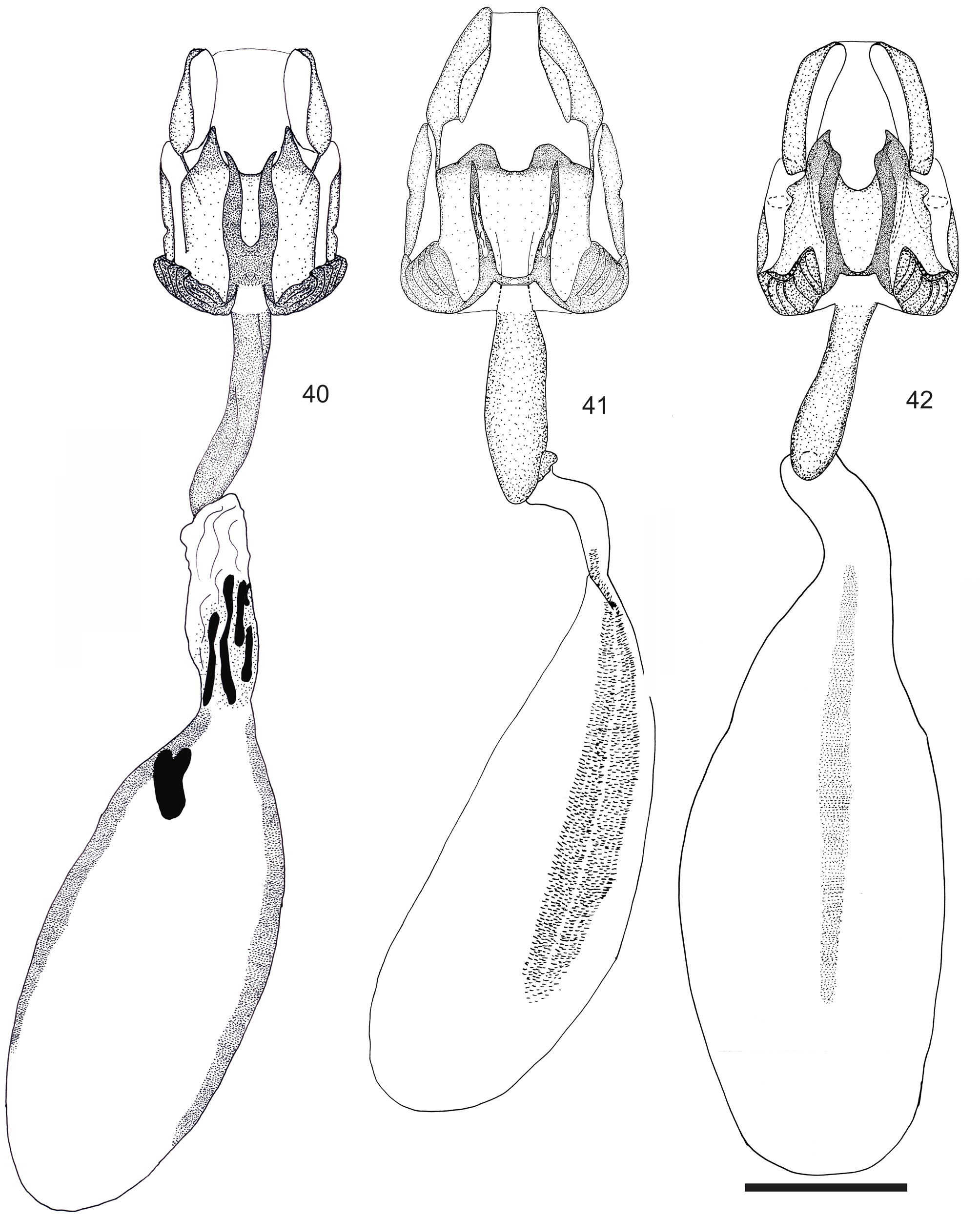
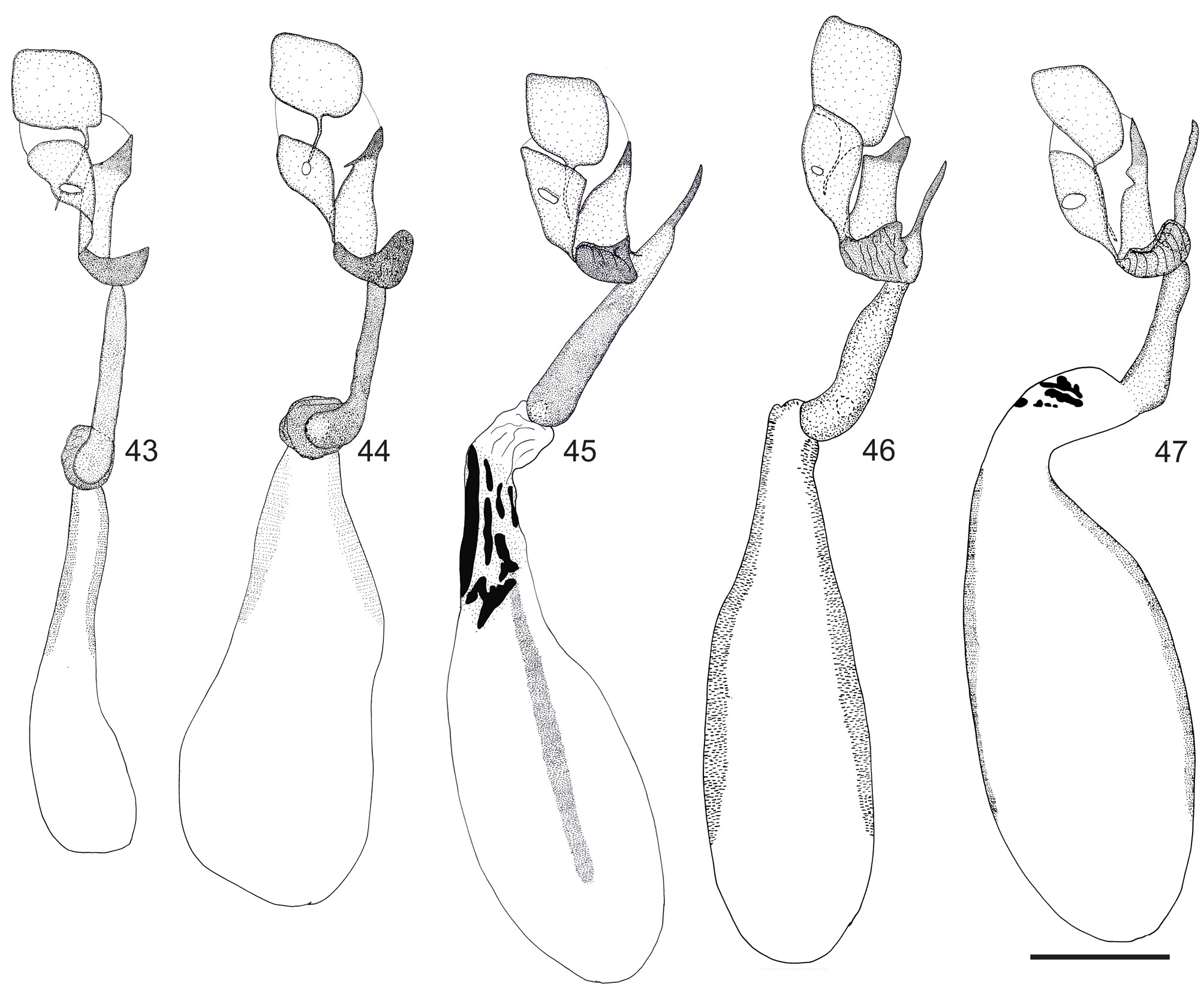
Diagnosis. Rufocumbre lucasi sp. nov. is distinguished from all the other species of Rufocumbre by the yellow costal area of the ventral forewing and basal and postdiscal bands of the ventral hind wing ( Figs 18, 20 View FIGURES 1–20 ); uncus widely bifid, with the arms thick, divergent and with a rounded tip; distal spine of the harpe short and pointed, never extending beyond the distal margin of ampulla ( Fig. 37 View FIGURES 35–37 ); lamella antevaginalis widely bifid, with broad and convergent projections, having the tips abruptly narrowed ( Fig. 42 View FIGURES 40–42 ).
Description. Forewing length: Male 14.8-16.8 mm, female 15.5-18 mm.
Forewing dorsal ( Figs 17, 19 View FIGURES 1–20 , 25 View FIGURES 21–25 ): brand with smooth margins; superior and inferior projections narrowed.
Forewing ventral ( Figs 18, 20 View FIGURES 1–20 ): subapical hyaline spots partially covered by yellow scales.
Hind wing ventral ( Figs 18, 20 View FIGURES 1–20 ): basal and postdiscal bands yellow, without gray to purple scales on the basal band; discal band narrow, proximally inclined near to costal margin.
Male genitalia ( Fig. 37 View FIGURES 35–37 ): Fenestra semicircular; uncus widely bifid, arms thick and divergent, tips rounded; ampulla largely projected distally, proximal margin with a broad and rounded projection, and a short dorsal concavity close to the distal margin; distal spine of harpe short and pointed, never exceeding the distal margin of ampulla; aedeagus distal end with unequal projections, cornutus present.
Female genitalia ( Figs 42 View FIGURES 40–42 , 47 View FIGURES 43–47 ): Lamella antevaginalis widely bifid from base, with broad and convergent projections, tips abruptly narrowed; lamella postvaginalis with the distal margin bifid, having two long, large, pointed and convergent projections; ductus bursae smooth at the insertion with the corpus bursae; corpus bursae with two long and large lines of signa, extending for almost all its extension.
Comments. Rufocumbre lucasi sp. nov. was never included in previous studies. The typical yellow basal and postdiscal bands of the hind wing of R. lucasi sp. nov. is unique among the species of Rufocumbre gen. nov. This color pattern, however, is shared with a single female of R. eberti from Serra da Maravilha, Senhor do Bonfim, Bahia ( Zacca & Bravo 2012). Thus, the analysis of the genitalia for both sexes of these species is required to ensure their identity. While dissection of the females of these species is necessary to distinguish them properly, dissection is not required to distinguish males of R. lucasi sp. nov. from those of R. eberti , since removing scales of the tip of the abdomen is sufficient to see whether the distal projection of the harpe exceeds the distal margin of the ampulla ( R. eberti , Fig. 35 View FIGURES 35–37 ) or not ( R. lucasi sp. nov., Fig. 37 View FIGURES 35–37 ).
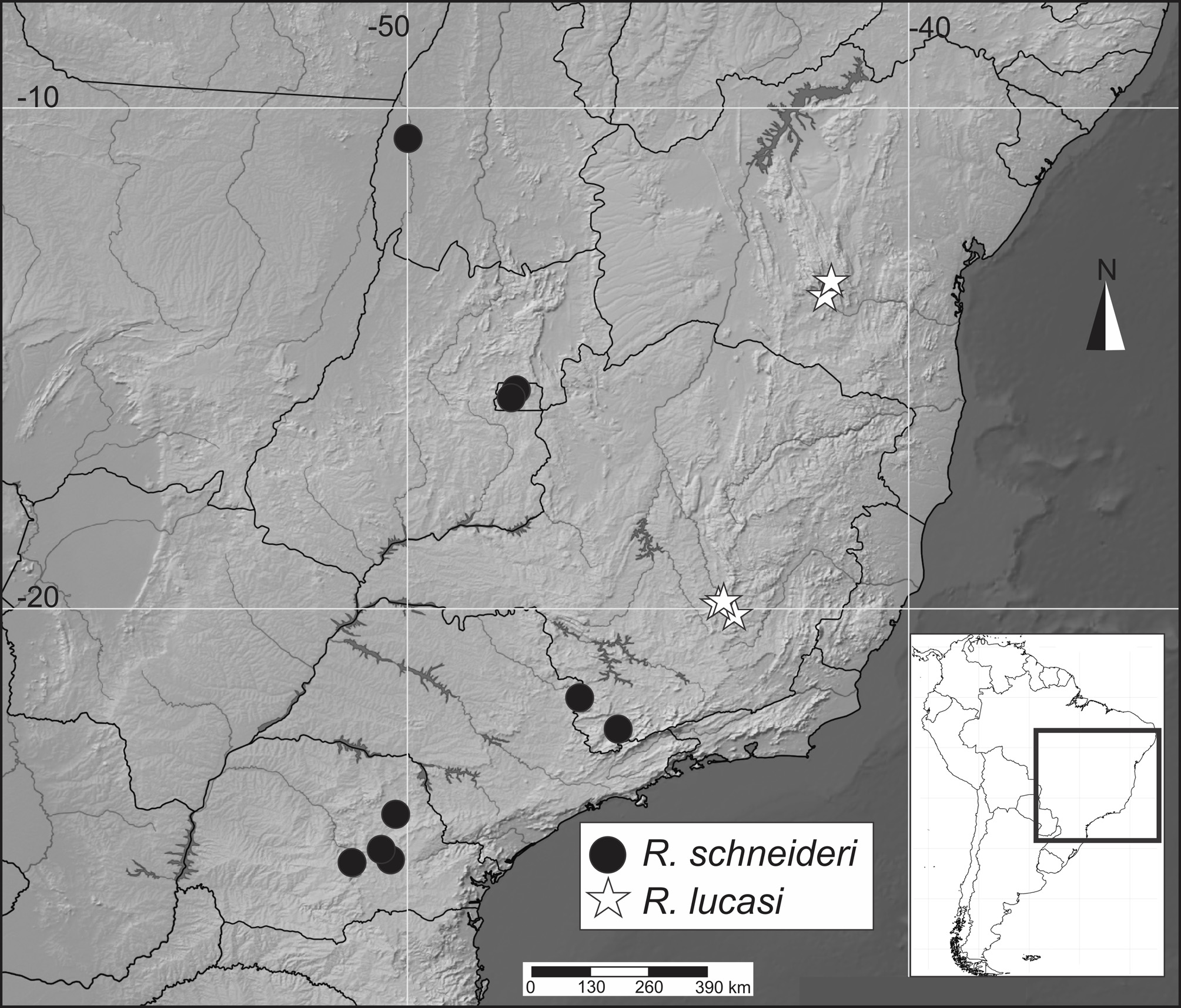
Geographical distribution and phenology. Rufocumbre lucasi sp. nov. is only known from areas throughout Serra do Espinhaço, in the states of Minas Gerais and Bahia, between 850 and 1600 m. Specimens were collected in January, March, August and September.
Etymology. This species is dedicated in memory of the late Lucas Caminha Beltrami, a student and friend who started the work with the genus Cumbre .
Examined material. The holotype male has the following labels: / HOLOTYPUS / 12-III-1999 [MORRO DO] PAI INÁCIO[,] PALMEIRAS , BAHIA [, BRAZIL,] 850m, MIELKE LEG. / OM 50.193 / HOLOTYPUS Rufocumbre lucasi Dolibaina, Mielke & Casagrande det. 2017/. DZUP.
The allotype female has the following labels: / ALLOTYPUS / BRASIL, MINAS GERAIS, CATAS ALTAS, CARAÇA, 16-18-IX-2006, 1300 m MIELKE & CASAGRANDE LEG. / DZ 15.584 / BC-DZ / ALLOTYPUS Rufocumbre lucasi Dolibaina, Mielke & Casagrande det. 2017/. DZUP.
Paratypes: BRAZIL – Bahia: Lençóis (Morro do Pai Inácio), 850 m, 10.III.1999, Mielke leg. 2 males (OM 50.009, OM 50.016*) (OM), 11.III.1999, Mielke leg. 1 male (OM 50.186) (OM), 12.III.1999, Mielke leg. 1 male and 1 female (OM 50.200*, OM 50.179*) (OM); Rio de Contas, 1400-1600 m, 28.I.2005, Mielke & Casagrande leg. 1 female (DZ 15.346*) (DZUP), (Mato Grosso), 1500 m, 29.I.2005, Mielke & Casagrande leg. 1 female (DZ 16.311) (DZUP), (Pico das Almas), 1450-1600 m, 30.I.2007, Mielke & Casagrande leg. 1 female (DZ 8.750*) (DZUP). Minas Gerais: Congonhas (Rio de Janeiro – Belo Horizonte Road, Km 344), 10-12.IV.1973, Callaghan leg. 3 males (MGCL); Nova Lima (Rio de Janeiro – Belo Horizonte Road, Km 432), 1400 m, 1.IV.1975, Callaghan leg. 1 male (MGCL).
Discussion
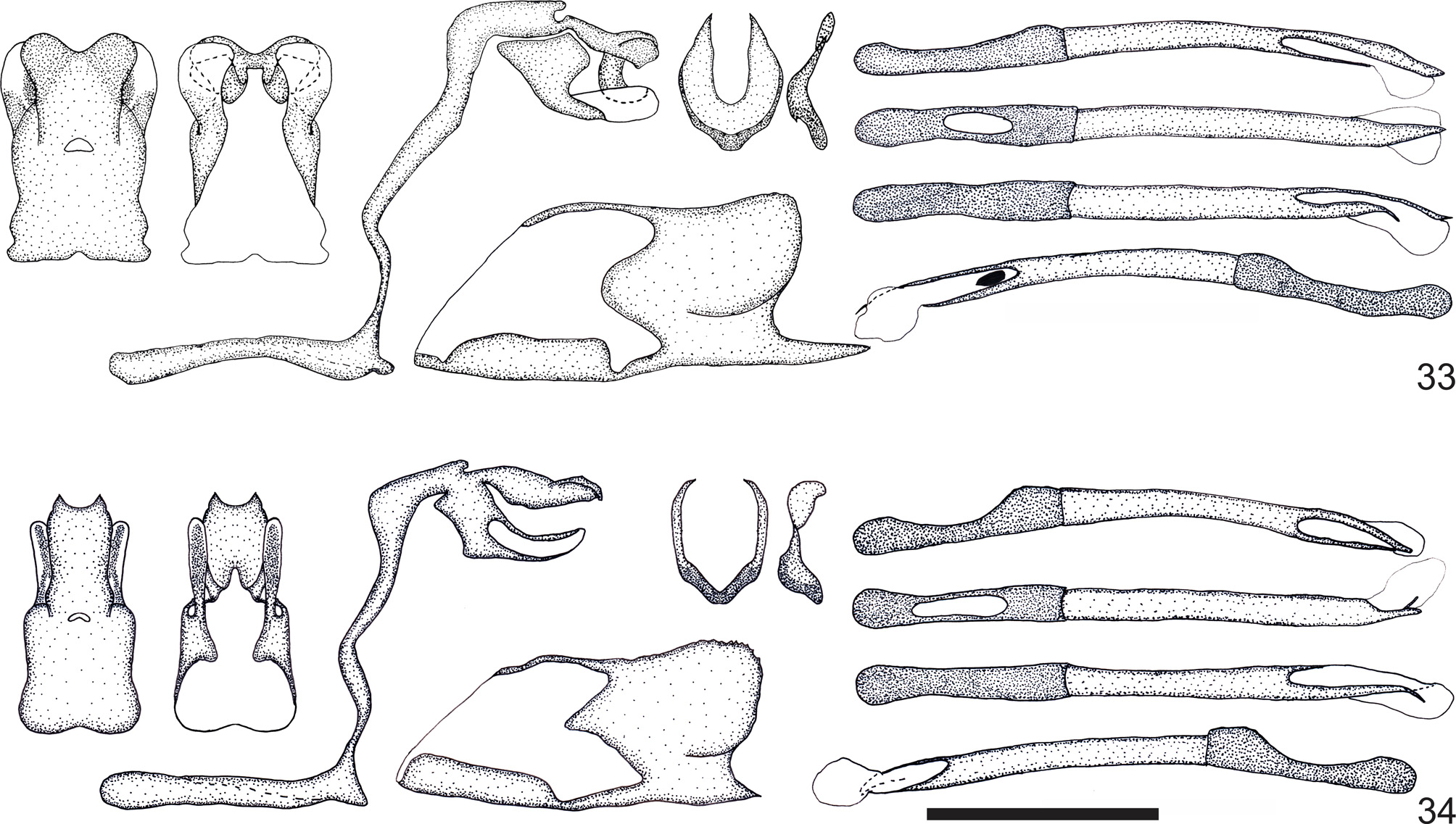
Two previously recognized taxa included in Rufocumbre were not seen by Evans (1955) as belonging to a distinct genus. In their revision of Cumbre, Dolibaina et al. (2014) pointed out that the subspecific taxon eberti belonged neither to Cumbre nor to any other known genus, and therefore required a new one. The reasons used by Evans (1955) to include R. celioi (misinterpreted by him as C. belli belli ) and R. eberti with the remaining species of Cumbre are not clear. One of the characters he used to circumscribe Cumbre was “uncus not divided” ( Evans 1955, p. 169). However, contrary to his written description of the genus, his schematic and partial illustration of the male genitalia of R. celioi ( Evans 1955, plate 65, J.41 CUMBRE 3. belli ) shows the uncus clearly bifid. In addition, in the synoptic collection of Evans deposited in the NHMUK, there is a schematic illustration of the male genitalia of R. eberti (not included in his monograph of 1955), with the uncus even more bifid than in R. celioi . Just the evident differences in the uncus could be used to place R. celioi and R. eberti in a genus apart from Cumbre . Moreover, the differences in the morphology of the uncus between R. celioi ( Fig. 33 View FIGURES 33–34 ) and R. eberti ( Fig. 35 View FIGURES 35–37 ) could have been used by Evans (1955) to consider the latter a distinct species, rather than a subspecies of the former.
The position of Rufocumbre among the Moncini genera is uncertain. The first systematic hypothesis involving two species of what is now Rufocumbre was the classification of Evans (1955), which included them ( R. celioi and R. eberti ) in the “ Lerema subgroup” of the “ Apaustus group” with nine other genera. However, the “ Apaustus group” and all its subgroups are not monophyletic ( Warren et al. 2008, 2009, Carneiro et al. 2015a).
Recent phylogenies using molecular and morphological characters recovered Rufocumbre celioi as sister to Ginungagapus ranesus (Schaus, 1902) ( Warren et al. 2008, 2009), to Cumbre cumbre Schaus, 1902 ( Carneiro et al. 2015a), and to Pheraeus odilia argus (Draudt, 1923) ( Sahoo et al. 2016). Such disparity calls for more phylogenetic studies addressing specifically Moncini , including a broader sampling of genera.
The morphology of the male genitalia has historically been used in the establishment of genera in Hesperiidae ( Godman 1900, Evans 1955, Mielke 1968, 1980, Carneiro et al. 2015a). On the other hand, characters of the female genitalia are poorly explored in the systematics of the group. Turland et al. (2012) stated that the presence and length of the sclerotized area of the ductus bursae of the female genitalia (antrum auct.) is a character indicative of higher relationships among Moncini genera. The ductus bursae found in Rufocumbre , which is sclerotized, cylindrical and as long as the sterigma is similar to that of Troyus turneri Warren & Turland, 2012 . Also, the ostium bursae at the base of the sterigma, and the absence of the lamella antevaginalis are features found in R. celioi , R. emeryi and T. turneri , suggesting the latter as a lineage potentially related to Rufocumbre . Several other genera representing all subgroups of the “ Apaustus group” have sclerotized ductus bursae, but, with a usually flattened aspect, and with the length of the sclerotized portion of the ductus more than twice the length of the sterigma (see discussion in Turland et al. 2012), thus being different greatly from Rufocumbre and Troyus . However, aspects of the wing color pattern, male forewing androconia and the remaining structures of the genitalia of both sexes suggest that Troyus Warren & Turland, 2012 is a distinct genus, though related to Rufocumbre .
Finally, more attention should be directed to redefining the Moncini genera, especially since a significant number of them have been found to be para or polyphyletic ( Burns 1990, Dolibaina et al. 2014, 2015, Carneiro et al. 2015a). It is essential to recognize independent lineages and to establish monophyletic units in an effort to resolve the Moncini phylogeny.



| DZUP |
Brazil, Parana, Curitiba, Universidade Federal do Parana, Museu de Entomologia Pe. Jesus Santiago Moure |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.