
Echinoderes, Yamasaki & Neuhaus & George, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4387.3.8 |
|
publication LSID |
lsid:zoobank.org:pub:DA75D56E-22BB-487E-8BA3-42C5C98F9E53 |
|
DOI |
https://doi.org/10.5281/zenodo.5952907 |
|
persistent identifier |
https://treatment.plazi.org/id/03CD87E0-2447-FFD7-3D86-29770F09FC4F |
|
treatment provided by |
Plazi |
|
scientific name |
Echinoderes |
| status |
|
Echinoderes sp. 1
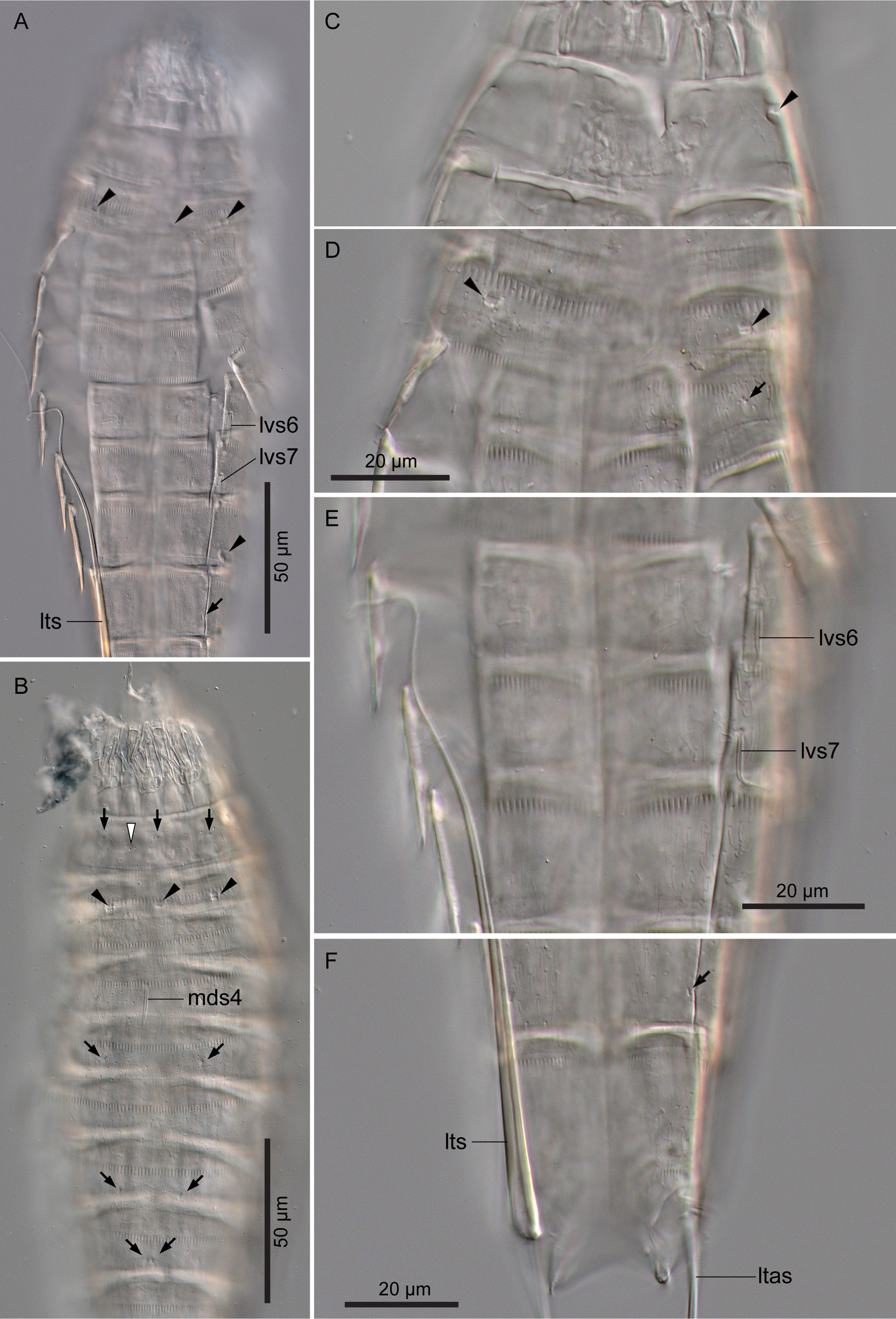
( Fig. 11 View FIGURE 11 ; Tables 6, 7)
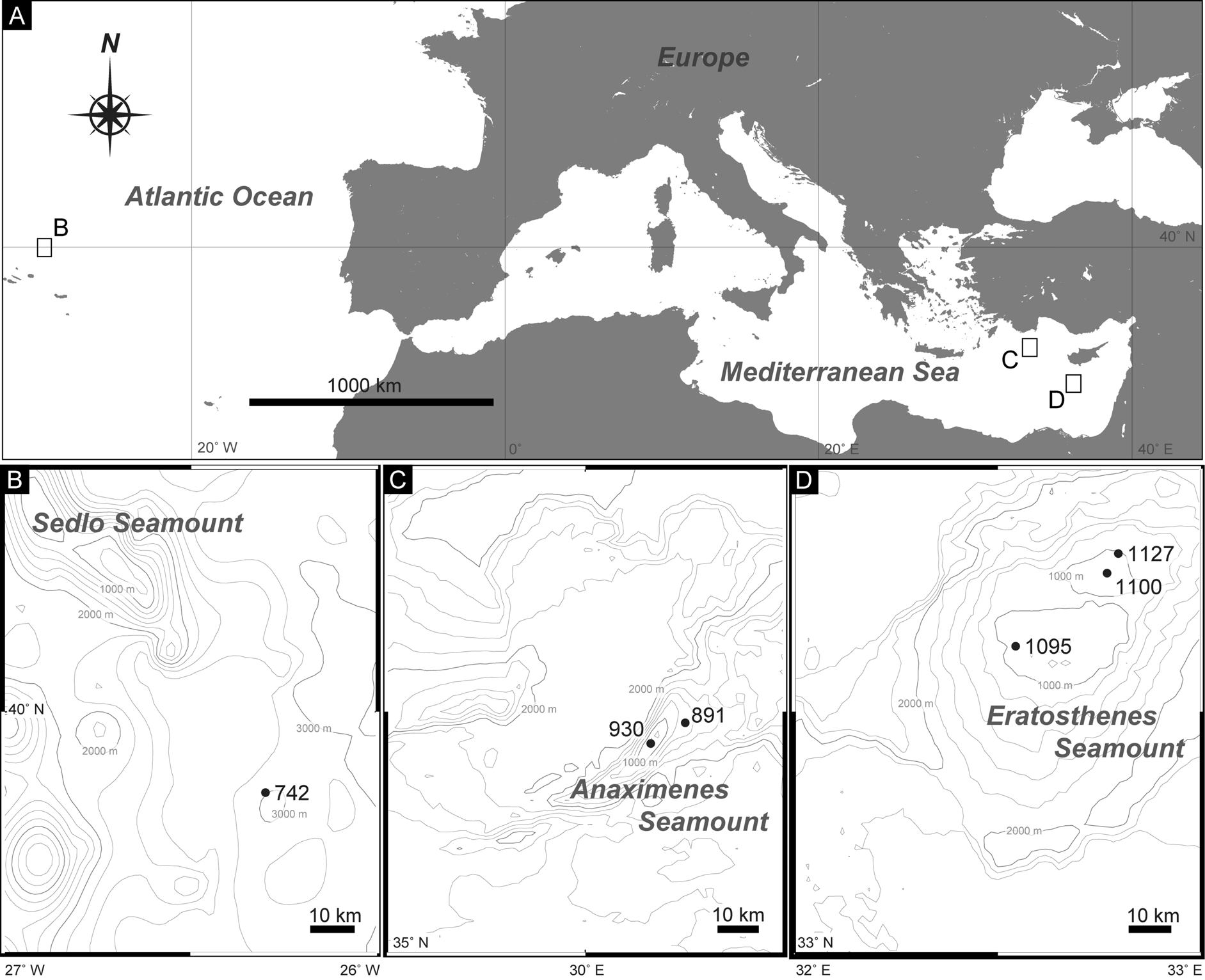
Material examined. One adult female ( ZMB 11593) collected at station 930 on Anaximenes Seamount, and one specimen of undetermined sex ( ZMB 11594) collected at station 891 on Anaximenes Seamount ( Fig. 1A, C View FIGURE1 ; Table 1). All specimens mounted as glycerol-paraffin slides on Cobb aluminum frames.
Brief description. Adult with head, neck, and eleven trunk segments. See Table 6 for measurements and Table 7 for positions of cuticular structures (sensory spots, glandular cell outlets, spines, and sieve plate).
Head consisting of retractable mouth cone and introvert. Mouth cone with inner oral styles and nine outer oral styles. Introvert composed of several rings of spinoscalids and one ring of trichoscalids. Exact number and arrangement of inner and outer oral styles and scalids not examined.
Neck with 16 placids. Midventral placid broadest. Remaining placids similar in size.
Segment 1 consisting of complete cuticular ring. This and following nine segments with thick pachycyclus at anterior margin. Pachycyclus interrupted middorsally in segments 2–10 as well as at tergosternal junctions in segments 3–10. Sensory spots located centrally in subdorsal and laterodorsal position ( Fig. 11B View FIGURE 11 ). Type-1 glandular cell outlets situated in middorsal and lateroventral position ( Fig. 11B View FIGURE 11 ). Type-2 glandular cell outlets present in midlateral position ( Fig. 11C View FIGURE 11 ). Posterior part of this and following nine segments with primary pectinate fringe ( Fig. 11D View FIGURE 11 ). Pectinate fringe teeth of primary pectinate fringes on this segment broader than those on following nine segments ( Fig. 11D View FIGURE 11 ).
Segment 2 with complete cuticular ring as segment 1. Sensory spots present in middorsal and ventromedial position. Type-1 glandular cell outlet in middorsal position. Type-2 glandular cell outlets in subdorsal, laterodorsal, sublateral, and ventrolateral position ( Fig. 11A, B, D View FIGURE 11 ).
Segment 3 and following eight segments consisting of one tergal and two sternal plates. Sensory spots present subdorsally and sublaterally ( Fig. 11D View FIGURE 11 ). Type-1 glandular cell outlets situated in middorsal position.
Segment 4 with middorsal acicular spine ( Fig. 11B View FIGURE 11 ). Sensory spots in subdorsal position. Type-1 glandular cell outlets present paradorsal and ventromedial position.
Segment 5 with sensory spots in subdorsal, midlateral, and ventromedial position ( Fig. 11B View FIGURE 11 ). Type-1 glandular cell outlets present in middorsal and ventromedial position. Type-2 glandular cell outlets present in lateral accessory position.
Segment 6 with lateroventral acicular spines ( Fig. 11A, E View FIGURE 11 ). Sensory spots present in paradorsal, midlateral, and ventromedial position. Type-1 glandular cell outlets present paradorsally and ventromedially.
Segment 7 with lateroventral acicular spines ( Fig. 11A, E View FIGURE 11 ). Sensory spots present in subdorsal, midlateral, and ventromedial position ( Fig. 11B View FIGURE 11 ). Type-1 glandular cell outlets present in middorsal and ventromedial position.
Segment 8 with paradorsal and laterodorsal sensory spots ( Fig. 11B View FIGURE 11 ). Type-1 glandular cell outlets present in paradorsal and ventromedial position. Type-2 glandular cell outlets situated in sublateral position ( Fig. 11A View FIGURE 11 ).
Segment 9 with paradorsal, laterodorsal, and ventrolateral sensory spots ( Fig. 11A, F View FIGURE 11 ). Type-1 glandular cell outlets present in paradorsal and ventromedial position. Small rounded sieve plates (nephridial openings) present in sublateral position.
Segment 10 with subdorsal and ventrolateral sensory spots. Two type-1 glandular cell outlets aligned middorsally. Additional pair of type-1 glandular cell outlets present in ventromedial position.
Segment 11 with lateral terminal spines ( Fig. 11A, F View FIGURE 11 ). Lateral terminal accessory spines present in female specimen ( Fig. 11F View FIGURE 11 ). Tergal plate extending laterally and forming tergal extensions.
Remarks. Echinoderes sp. 1 is similar to E. unispinosus sp. nov. in having acicular spines only in middorsal position on segment 4 and in lateroventral position on segments 6 and 7. There is no other species with such a spine formula. In addition, the positions of type-2 glandular cell outlets are completely identical in the two species. These similarities suggest the close relationship between the two species.
Despite these similarities, we did not assign Echinoderes sp. 1 to Echinoderes unispinosus sp. nov. but assume it to represent another undescribed species because of several morphological differences between the two species: Echinoderes sp. 1 is smaller in trunk length than E. unispinosus sp. nov. (219 µm in Echinoderes sp. 1, whereas 265–305 µm in E. unispinosus sp. nov.), in Echinoderes sp. 1 the pectinate fringe teeth of the primary pectinate fringe on segment 1 are broader than those on the other segments of this species, whereas the width of the pectinate fringe teeth is similar on all segments in E. unispinosus sp. nov. (compare Fig. 9A View FIGURE9 and Fig. 11D View FIGURE 11 ), sublateral sensory spots on segment 3 and subdorsal sensory spots on segment 4 are present in Echinoderes sp. 1, whereas such spots are absent in E. unispinosus sp. nov., and ventromedial sensory spots on segment 8 are absent in Echinoderes sp. 1, whereas such spots are present in E. unispinosus sp. nov. However, the difference in size and the presence/absence of sensory spots may just originate from variation in one and the same species. Such variations were revealed in other kinorhynch species, e.g., Campyloderes cf. vanhoeffeni Zelinka, 1913, Cateria gerlachi Higgins, 1968 , Cateria styx Gerlach, 1956 , Centroderes barbanigra Neuhaus et al., 2014 , Ce. drakei Neuhaus et al., 2014, Ce. readae Neuhaus et al., 2014, Ce. spinosus (Reinhard, 1881) (Neuhaus et al. 2013, 2014; Neuhaus & Sørensen 2013; Neuhaus & Kegel 2015). The variation within these species was investigated based on a larger amount of specimens, whereas a very limited amount of specimens of E. unispinosus sp. nov. and of Echinoderes sp. 1 are available for the current study. Furthermore, all specimens of Echinoderes sp. 1 in this study broke during the preparation. Thus, we hesitate to formally describe the species as a new species in this paper. We hope the species will be named as a new species when further material will become available.

| ZMB |
Museum f�r Naturkunde Berlin (Zoological Collections) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.