
Wilcoxia James, 1941
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4695.5.1 |
|
publication LSID |
lsid:zoobank.org:pub:A6844BF7-1B08-40D8-AB38-37F1E93B434C |
|
persistent identifier |
https://treatment.plazi.org/id/03CD87EE-0814-F049-FF48-F9C3FBB2909E |
|
treatment provided by |
Plazi |
|
scientific name |
Wilcoxia James |
| status |
|
Wilcoxia James View in CoL View at ENA
Wilcoxia James, 1941 View in CoL . Type species, by monotypy and original designation: W. cinerea James, 1941 View in CoL ; Wilcox & Martin 1957: 2 [key to related genera]; Hull 1962 a, b: 122 [in key to genera of Stenopogonini ], Figs 82 (antenna), 585 (wing), 980 (frontal view of head); Martin & Wilcox 1965: 385 [catalog of Nearctic Diptera ]; Wilcox 1972: 43; Wood 1981: 569 [key to Nearctic genera]; Papavero 1973: 248 [in list of genera in subfamily Stenopogoninae View in CoL ]; Artigas & Papavero 1991 a: 142 [in list of tribes and genera of American Stenopogoninae View in CoL ]; Artigas & Papavero 1991 b: 57 [in key to genera of Stenopogoninae View in CoL : Cyrtopogonini ]; Poole & Gentili 1996: 64.
Notes. James (1941) did not provide a description of his new genus, but a relatively detailed description was given for W. cinerea View in CoL , then the only included species. Hull’s (1962 a) description of the genus was more complete than that of James (1941) but was probably also based on only W. cinerea View in CoL . However, in the captions for the four figures de- picting a species of Wilcoxia in Hull (1962 b) View in CoL , only one of these includes W. cinerea View in CoL ; the other three merely list the illustrated specimen as Wilcoxia View in CoL sp.
Poinar (1992) illustrated a robber fly from Dominican amber (Oligocene/Miocene), called a Wilcoxia in the caption. This has led to the genus being included in the Neotropical region (e.g. Papavero 2009, as Wilcoxia sp.). The specimen on which this identification was based is deposited in the American Museum of Natural History (Poinar, pers. comm.). According to Eric Fisher (pers. comm.) who has examined this amber inclusion, a preliminary identification of “ Wilcoxius sp.” by him was misinterpreted by Poinar and given instead as “ Wilcoxia sp.”. Wilcoxius is a Neotropical genus in the Asilinae.
Species groups.—As discussed below, when Wilcox (1972) described four new species of Wilcoxia , the genus became structurally heterogeneous; however, the presence of the distinct mesotibial spine seems to have been considered a diagnostic feature for the genus. Though this structure is found in all species currently in Wilcoxia (including the three new species described herein), we have observed other characters that seem to divide the eight species into two species groups. We have not established new subgenera or a new genus (for the martinorum group), but rather suggest that this might be a necessary action in the future based on further study including phylogenetic analysis. Structural features which divide the species into the two species groups ( cinerea and martinorum ): relative length and angle of projection of dorsocentral hairs and setae on the scutum; number and angle of projection of marginal setae on scutellum; “crinkliness” of katatergite setae; pattern of pollinosity on abdominal tergites; and color of legs. Another observation included here is the tendency for specimens of the two different species groups to be collected either relatively early or late in the year. No Wilcoxia collections are known from March to May, and specimens have been collected on only one day in both February and June. The four species in the cinerea group were collected from June to October (only a single specimen in October), while the four species in the martinorum group were collected from October to February ( Table 1 View TABLE 1 ).
Description. Small robber flies, body length, excluding antennae from 4.4–9.9 mm; wing lengths from 4.3–6.9 mm. Head ( Figs 17–25 View FIGURES 17–25 ). Width 1.5–1.7 times eye height; face pollinose, very short, in profile at, or slightly beyond eye margin; facial gibbosity only slightly developed; sides of face very slightly to moderately converging toward antennae; frons pollinose, subparallel-sided to moderately diverging towards vertex; compound eyes relatively large, occupying almost entire height of head; posterior margins evenly arcuate or slightly angulate (esp. so in martinorum group); anterior facets distinctly enlarged; ocellar tubercle very slightly to moderately elevated, bearing short and longer setae; median ocellus larger than lateral ocelli; occiput densely pollinose, flat to moderately convex, dorsally with long, postocular bristles, ventrally bearing occipital hairs of various lengths and densities (the latter esp. well developed in martinorum group, Figs 21–25 View FIGURES 17–25 ); mystax variable, occupying lower half of face in some species, in others occupying entire face, especially long and dense in martinorum group species (e.g., Figs 21, 23 View FIGURES 17–25 ); face above mystax with smaller, appressed hairs; proboscis laterally slightly compressed, blunt apically; palpi 2-segmented, distal palpomere distinctly longer than proximal; antennae inserted slightly above middle of head, antennal bases not on raised area; insertions approximate, separated by scape width or less; scape and pedicel short, subequal in length, with various number of short and/or long setae; scape subfiliform, parallel-sided; pedicel submoniliform, slightly to moderately widened distally; postpedicel elongate, longer than scape and pedicel combined, parallelsided for at least ½ length, then slightly to moderately tapered apically; style 2-segmented, first much shorter than second, but of equal widths; second stylomere with short slender spine; antennae pollinose, some species with bare scape (sometimes just the medial surface).
Thorax. Entire surface pollinose; ante- and postpronotum with variable number of setae; scutum moderately convex; most species with divided median stripe between 2 rows of acrostichal bristles; surface uniformly pollinose, some species with one or more elliptical bare spots or spots with contrastingly colored tomentum, on each side of midline; vestiture of scutum various, some species with long presutural setae, others with very short setae, with any longer setae restricted to postsutural area of scutum; notopleural, supraalar and postalar bristles present, variable in number (from 1–3); anepisternum and katepisternum without setae; katatergite bearing variable number of long setae, apices either “crinkly” ( martinorum group, e.g. Fig. 43 View FIGURES 41–46 ) or not ( cinerea group, e.g. Fig. 44 View FIGURES 41–46 ); anterior surface of procoxae and lateral surfaces of meso- and metacoxae with elongate hairs; scutellum slightly convex, entire surface pollinose in some species (e.g., Fig. 41 View FIGURES 41–46 ), in others, with posterior margin shining, non-pollinose (e.g., Fig. 42 View FIGURES 41–46 ); disc of scutellum with or without scattered, short hairs; posterior margin with longer setae, the angle compared to plane of scutellum ranging from nearly parallel to perpendicular, setae either parallel to each other, or distinctly converging medially.
Legs relatively slender, similar in proportions among pairs; color uniformly dark (piceous to black) ( martinorum group, Figs 9–16 View FIGURES 9–12 View FIGURES 13–16 ) or with contrasting dark and yellow areas ( cinerea group, Figs 1–8 View FIGURES 1–4 View FIGURES 5–8 ); hind leg moderately longer than anterior two pairs; peg-like protuberance present on anterior face of metacoxae; femora relatively slen- der, elongate, variously swollen in proximal half, or subparallel; surface of femora bare ( cinerea group), pollinose, or a combination of the two (spp. in martinorum group), covered with scattered, appressed short hairs and longer, erect setae; dorsal apex of femora with or without one to several stout setae/bristles; tibiae slender, straight, subequal in length to femora; surface with combination of short, appressed hairs and longer setae/bristles, similar on all legs (male W. apache with anterior tibial hairs much longer than on other legs); apex of front tibiae without or with ( W. painteri only) short, slightly sigmoidal spine ( Fig. 45 View FIGURES 41–46 ); ventral apex of middle tibiae with stout spine/bristle ( Fig. 46 View FIGURES 41–46 ), in most species darker than regular tibial bristles; tarsi 5-segmented, relatively slender, more robust in some species, middle tarsomeres submoniliform; length of tarsus subequal in length to tibia; hind tarsus, at least in male, more robust than front or middle tarsus; brush of setae on first tarsomere somewhat denser on hind tarsus; length of tarsomere 1 subequal in length to 2 + 3; apex of each tarsomere with circlet of relatively long, stout bristles; tarsal claws fine and sharp, widest basally, then abruptly narrowed, distinctly curved apically; pulvilli long, slender, almost as long as claws; empodium slender, about half length of tarsal claw.
Wing ( Figs 38–40 View FIGURES 38–40 ) immaculate, hyaline to moderately darkly infuscated brown, color slightly sexually dimorphic (darker in males) in some species of cinerea group; costa (C) complete around posterior margin to wing base, except in W. forbesi , in which it stops near junction of CuA 2 + A 1 ( Fig. 38 View FIGURES 38–40 ); vein R 2+3 ending in C, apex very slightly directed forward before C; R 4 and R 5 only slightly arcuate; r-m crossvein beyond midlength of discal cell; apical portion of M 3 at approximately right angle to m-m crossvein; A 1 and CuA 2 meeting at margin or with slight gap; cell m 3 broadly open at wing margin; cell cu p closed or very narrowly open at wing margin; wing membrane either without conspicuous microtrichiae (esp. martinorum group, e.g. Fig. 40 View FIGURES 38–40 ) or with distal half to third with dense, conspicuous microtrichiae ( cinerea group, e.g. Fig. 39 View FIGURES 38–40 ).
Abdomen relatively slender, convex dorsally; first segment slightly narrower than width of mesonotum; segments wider than long or subequal in length and width (in W. pollinosa segments 2–4 distinctly longer than wide); males with 7 visible tergites; female with 8 visible tergites, 8 th about half length of 7 th; tergites densely pollinose with bare patches vaguely in “checkerboard” pattern (most martinorum group spp., Figs 30–33, 36–37 View FIGURES 30–37 ) or mostly bare (most cinerea group spp., Figs 26–29 View FIGURES 26–29 and female of W. forbesi from the martinorum group, Fig. 35 View FIGURES 30–37 ) with pollinosity restricted to narrow lateral and/or anterior bands; male and female W. pollinosa ( Fig. 29 View FIGURES 26–29 ) and male of W. forbesi ( Fig. 34 View FIGURES 30–37 ) with tergites uniformly pollinose; tergites dorsally with relatively short, sparse setae (relatively longer in spp. of martinorum group), laterally with longer setae, especially on tergites 1–3 (very long in some martinorum group spp.); sternites uniformly pollinose, 6–7 of females of some species bare, non-pollinose; surface of sternites with setae/hairs of various lengths, relatively inconspicuous in some species (especially W. pollinosa ), in others very long ( martinorum group), longer on sternites 1–3 than posterior sternites.
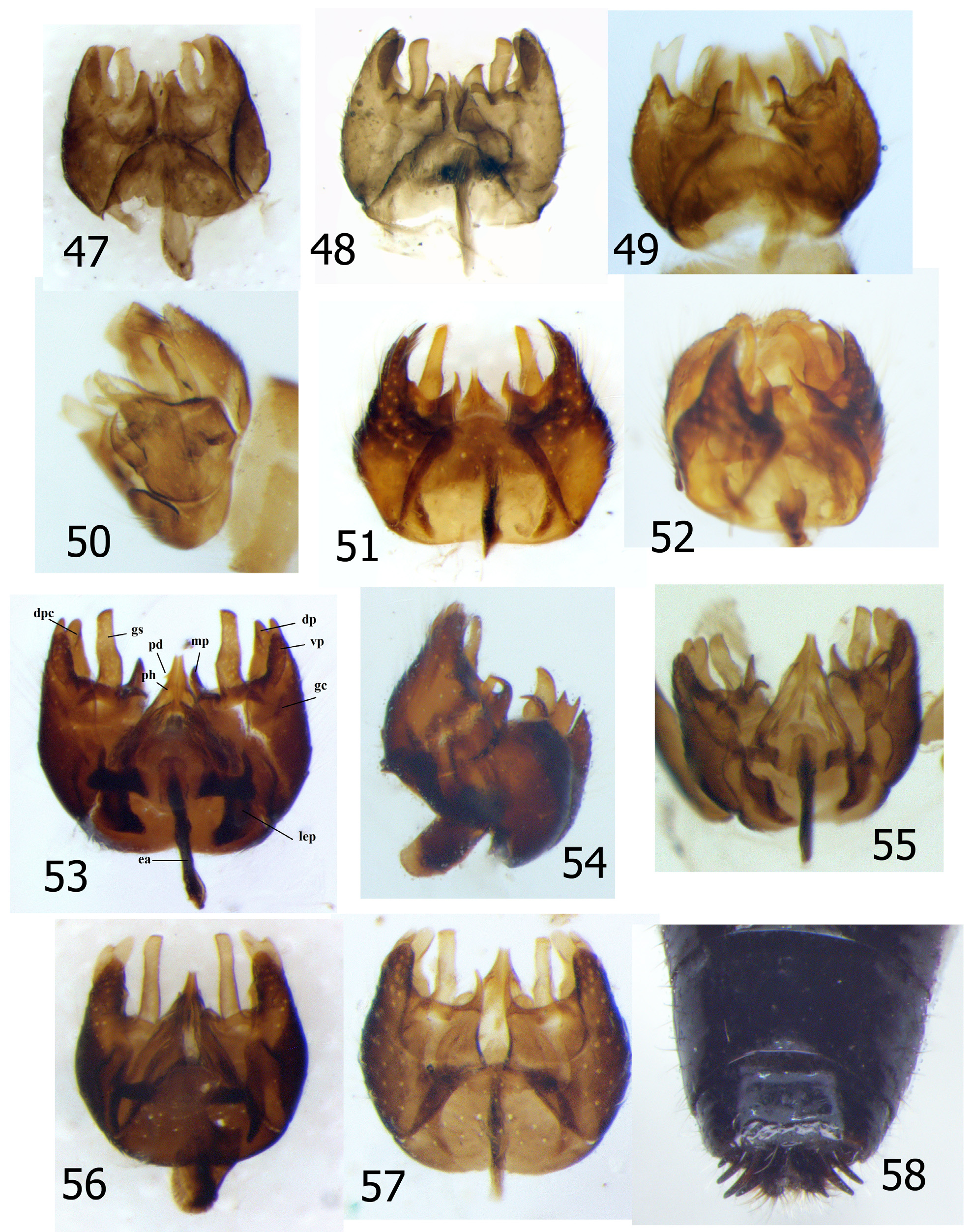
Male genitalia ( Figs 47–57 View FIGURES 47–58 ) relatively short, inconspicuous, rotated 90–180° in most specimens; genitalia somewhat oblique in lateral view, with dorsal structures protruding more posteriorly than ventral; hypandrium distinct, widest anteriorly, roughly triangular to subtrapezoidal in shape; epandrium divided into two separate lobes, attached posteriorly; cerci short, broadly rounded to subtruncate; gonopod well-developed, consisting of elongate, digitiform gonostylus ( Fig. 53 View FIGURES 47–58 , gs) and gonocoxite; gonocoxite with relatively robust dorsal ( Fig. 53 View FIGURES 47–58 , dp) and ventral processes ( Fig. 53 View FIGURES 47–58 , vp) laterally, and medial process ( Fig. 53 View FIGURES 47–58 , mp); dorsal process variously bifid (e.g., Figs 53–54 View FIGURES 47–58 ) or entire; medial process slender, indistinctly to very distinctly apically curved dorsad; phallus ( Fig. 53 View FIGURES 47–58 , ph) very wide basally, evenly or somewhat abruptly narrowed distally, with or without small denticle ( Fig. 53 View FIGURES 47–58 , pd) on lateral margins.
Female genitalia. Tergite 10 divided into two acanthophorites, each bearing 4–5 short, slightly curved spines ( Fig. 58 View FIGURES 47–58 ); spermathecae (based on Figs 37–38 View FIGURES 30–37 View FIGURES 38–40 in Artigas and Papavero 1991 b) three in number, very elongate, thread-like.
Natural history. Relevant biocontextual information is generally lacking for most species of Wilcoxia ; specific details are presented below under each species. The best-known species are those in the martinorum group (especially W. apache , new species, given its proximity to Portales, the authors’ town of residence), perhaps because of their tendency to rest on rocky substrates (e.g., Figs 68–72 View FIGURES 67–70 View FIGURES 71–76 ) and therefore their relative ease of observation and/or collection. In fact, all known prey associations for Wilcoxia are from the martinorum group: W. apache , new species, W. forbesi , new species, and W. martinorum . It is hypothesized that members of the cinerea group are more closely tied to vegetation and therefore their habits are more difficult to observe.











No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Stenopogoninae |
Wilcoxia James
| Pollock, Darren A. & Reichert, Lisa A. 2019 |
Wilcoxia
| Poole, R. W. & Gentili, P. 1996: 64 |
| Artigas, J. N. & Papavero, N. 1991: 142 |
| Artigas, J. N. & Papavero, N. 1991: 57 |
| Wood, G. C. 1981: 569 |
| Papavero, N. 1973: 248 |
| Wilcox, J. 1972: 43 |
| Martin, C. H. & Wilcox, J. 1965: 385 |
| Wilcox, J. & Martin, C. H. 1957: 2 |