
Hallodapini Van Duzee, 1916
|
publication ID |
https://doi.org/10.2478/aemnp-2019-0007 |
|
publication LSID |
lsid:zoobank.org:pub:027CE86F-9E75-44C3-A35E-E0C20BA4B693 |
|
DOI |
https://doi.org/10.5281/zenodo.5062794 |
|
persistent identifier |
https://treatment.plazi.org/id/03CDBD54-EC5D-FFFA-FC22-FDF3FA93FC03 |
|
treatment provided by |
Felipe |
|
scientific name |
Hallodapini Van Duzee, 1916 |
| status |
|
Tribe Hallodapini Van Duzee, 1916
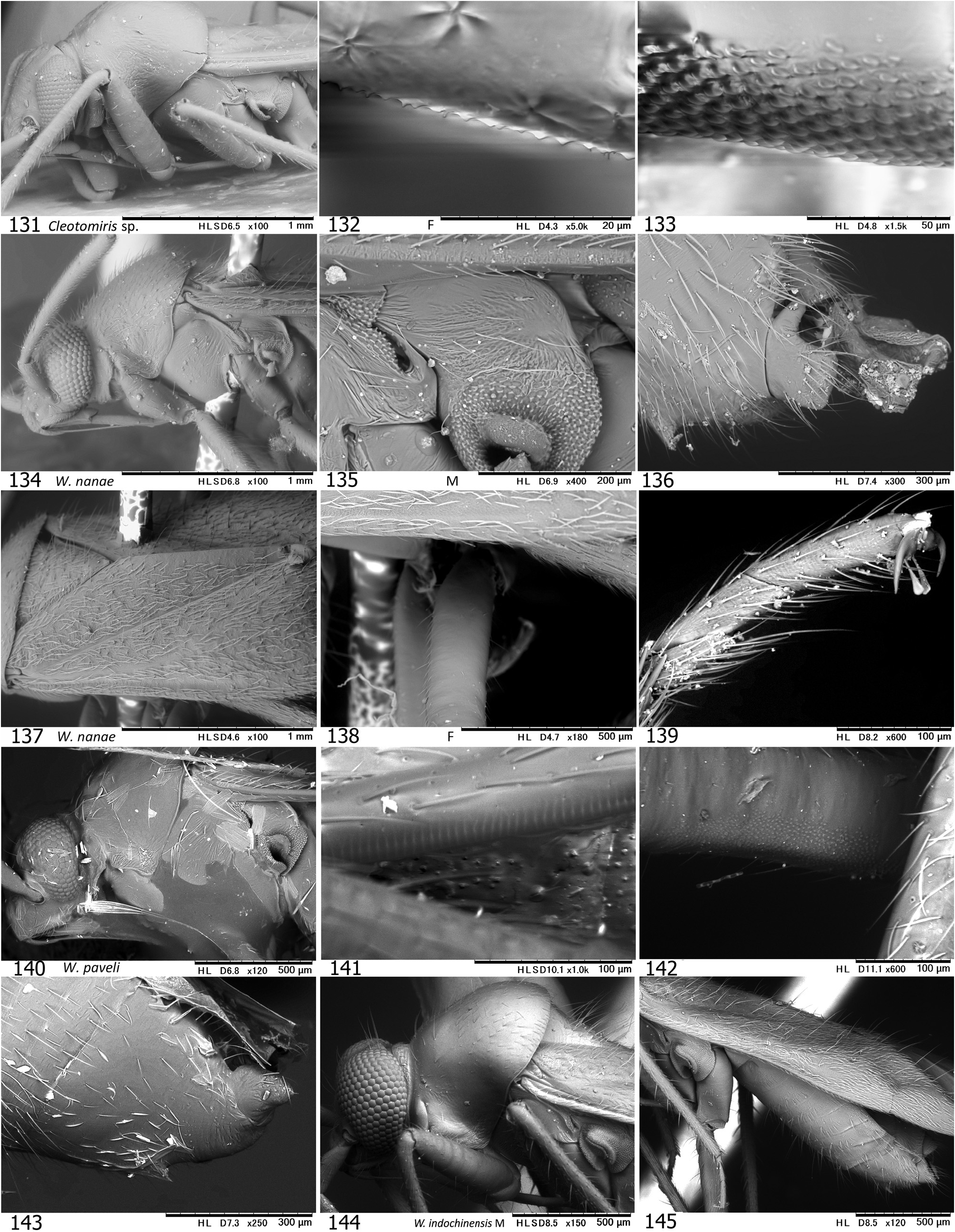
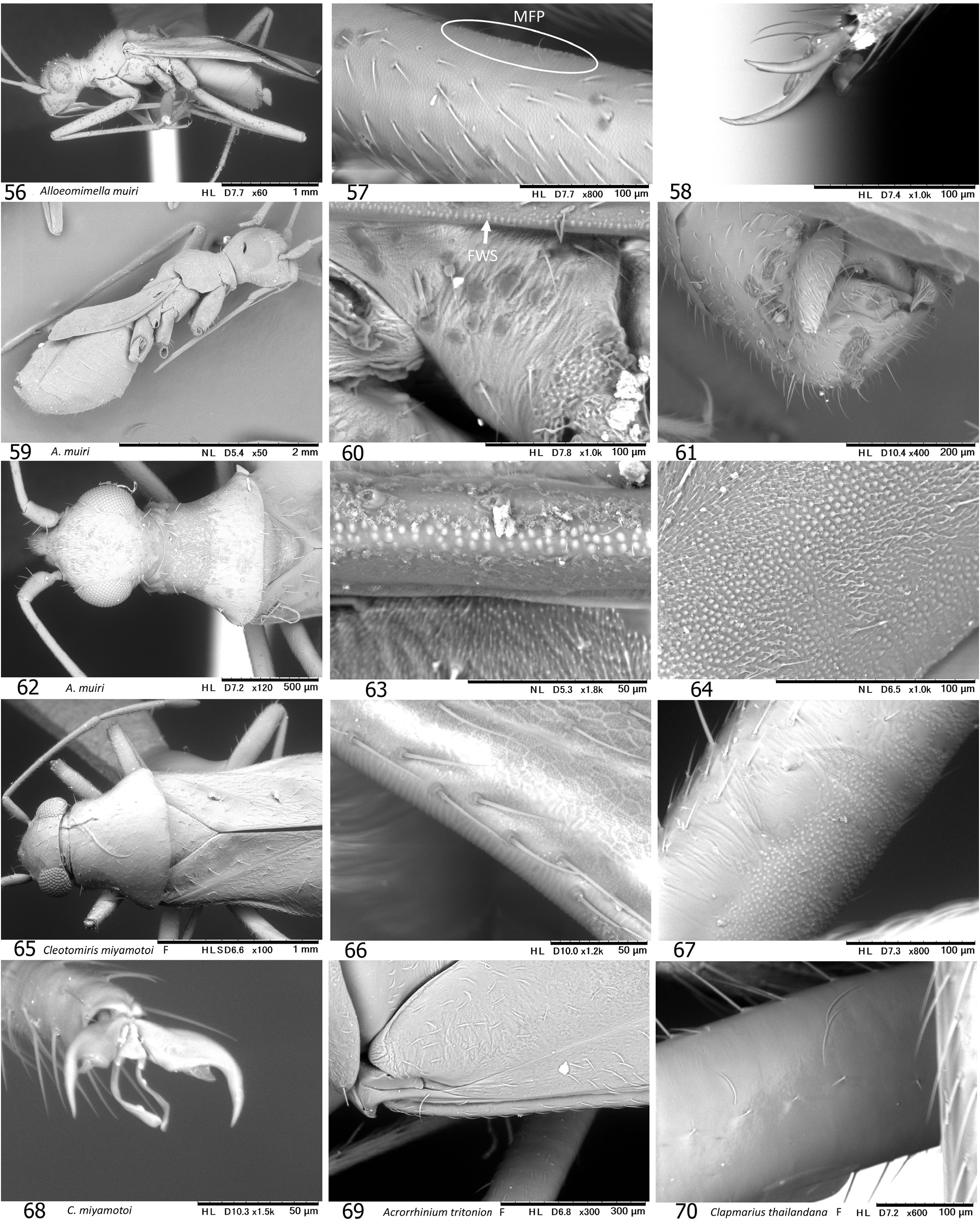
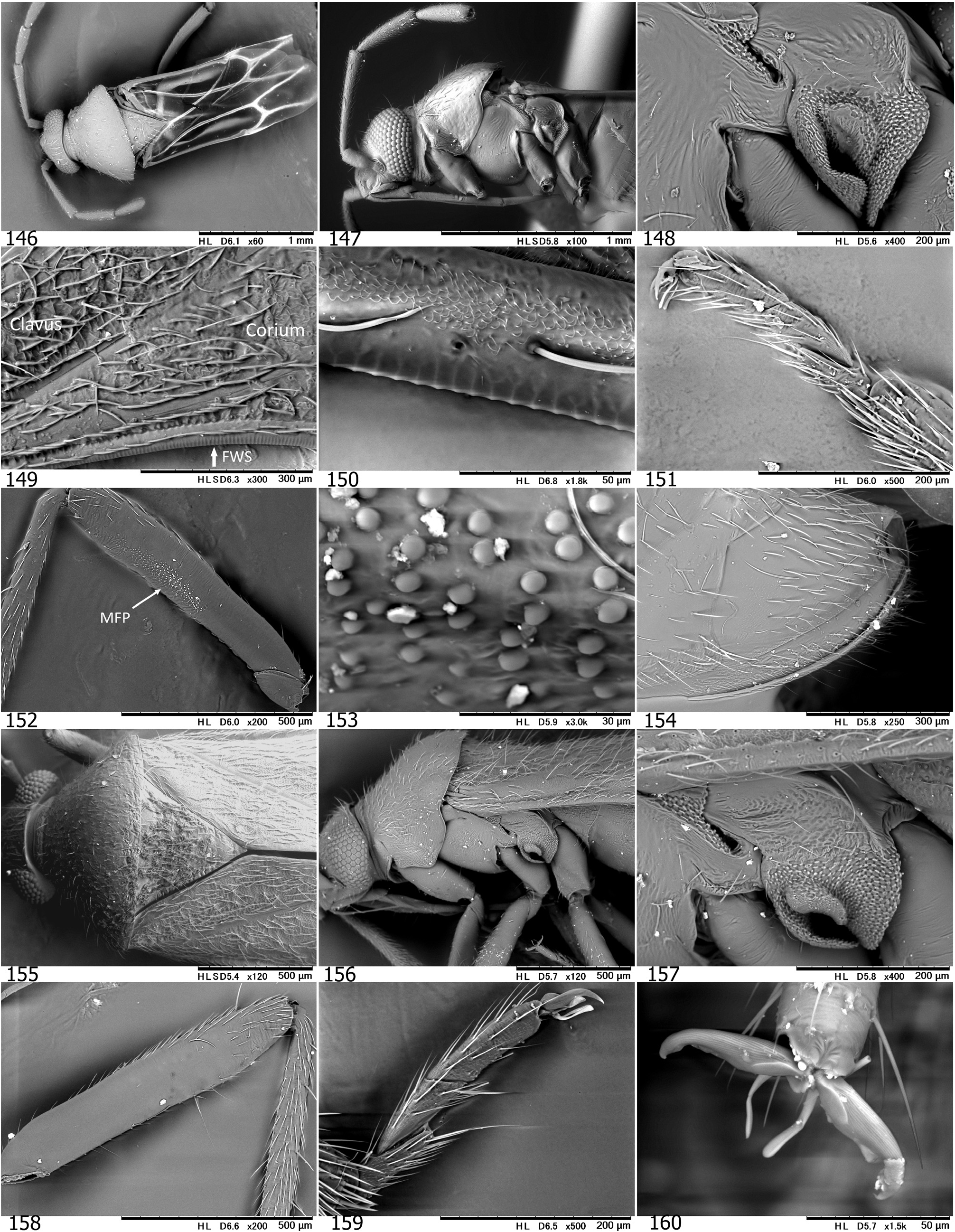
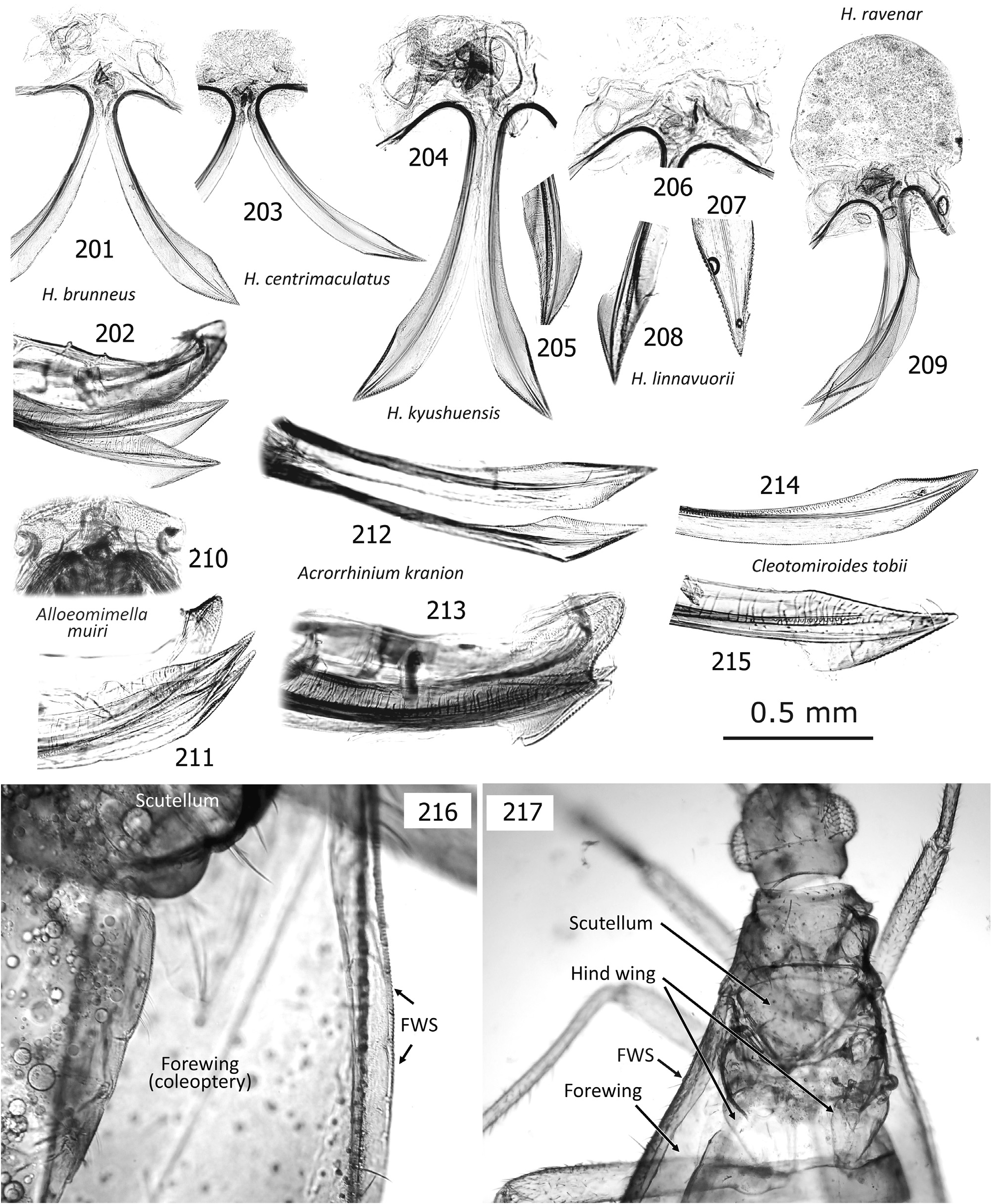
Diagnosis. Distinguished from any other tribes in the subfamily Phylinae by the following combination of characters: Body elongate to elongate oval, sometimes more slender (e.g., Podullahas Schuh, 1984 , Sohenus Distant, 1910 ) variable in size (2.2–7.0 mm), more or less antlike ( Figs 9–24 View Figs 9–18 View Figs 19–24 ), with simple vestiture, lacking scalelike setae; brachypterous form often present mostly in female (coleoptery or staphylinoidy sensu SCHUH & SLATER 1995); basic coloration brownish or reddish, without green tinge; eyes small; antenna uniformly long, almost linear and often thickened, as long as or longer than body; pronotum usually constricted anteriorly, with distinct, flat collar; margin of exocorium (FWS) minutely notched or serrate and dorsal surface of metafemur furnished with minute bumps (plectrum, MFP) forming a stridulatory device in six genera; legs generally long; left paramere with developed sensory lobe tightly holding phallotheca; right paramere usually reduced, tiny; endosoma elongate in most members, often sigmoid or coiled, with more or less elaborated apical part and thick-rimmed secondary gonopore; sclerotized ring ovoid to elongate oval, clearly rimmed.
Distribution. Nearly cosmopolitan but majority of species known from the Old World tropics, subtropics and warm temperate zones; no reliable record existing from Pacific islands or atolls; only 13 species in three genera, Cyrtopeltocoris Reuter, 1876 , Lapazphylus Carvalho & Costa, 1992 and Phoradendrepulus Polhemus & Polhemus, 1985 known in New World and Acrorrhinium spicatum (Distant, 1904) in Australian Region (SCHUH 2013). However, some New World genera treated by SCHUH & MENARD (2013) as incertae sedis in the Phylini could be true hallodapine members (K. L. Menard, pers. comm.).
Discussion. SCHUH (1984) proposed Auricillocorini composed of five Oriental genera, Auricillocoris Schuh, 1984 , Cleotomiris , Cleotomiroides , Wygomiris and Zaratus Distant, 1909 . Although this classification was followed by YASUNAGA (2012), the Auricillocorini is now regarded as a junior synonym of Hallodapini , involving 14 valid genera in Asia (cf. SCHUH 2013, SCHUH & MENARD 2013). Possession of the stridulatory device is currently confirmed in members of six Old World genera. In addition to Asian Alloeomimella new genus, Cleotomiris , Hallodapus , and Wygomiris ( Table 1 View Table 1 ), two Ethiopian genera, Laemocoris Reuter, 1879 and Trichophthalmocapsus Poppius, 1914 , were reported to have the lateral hemelytral margins (FWS) and dorsobasal surface of metafemora (MFP) modified to form a stridulatory mechanism ( SCHUH 1974).
We have successfully confirmed that Hallodapus centrimaculatus (approx. 2,100 Hz) and H. ravenar (800 Hz) produced sounds, using the stridulatory device (FWS+MFP) during courtship behavior ( Fig. 32 View Fig ). Nonetheless, our recent observations suggest that the presence of the stridulatory device may be homoplasious in some species of a certain genus. This character status is evidenced by closely related species of Wygomiris ; in W. indochinensis ( Fig. 145 View Figs 131–145 ) vs. W. paveli ( Figs 141–142 View Figs 131–145 ) and W. kaliyahae ( Fig. 58 View Figs 56–70 ) vs. W. phormictes ( Figs 150, 153 View Figs 146–160 ) – the former species lacks the device, whereas the latter possesses it (see Table 1 View Table 1 ).
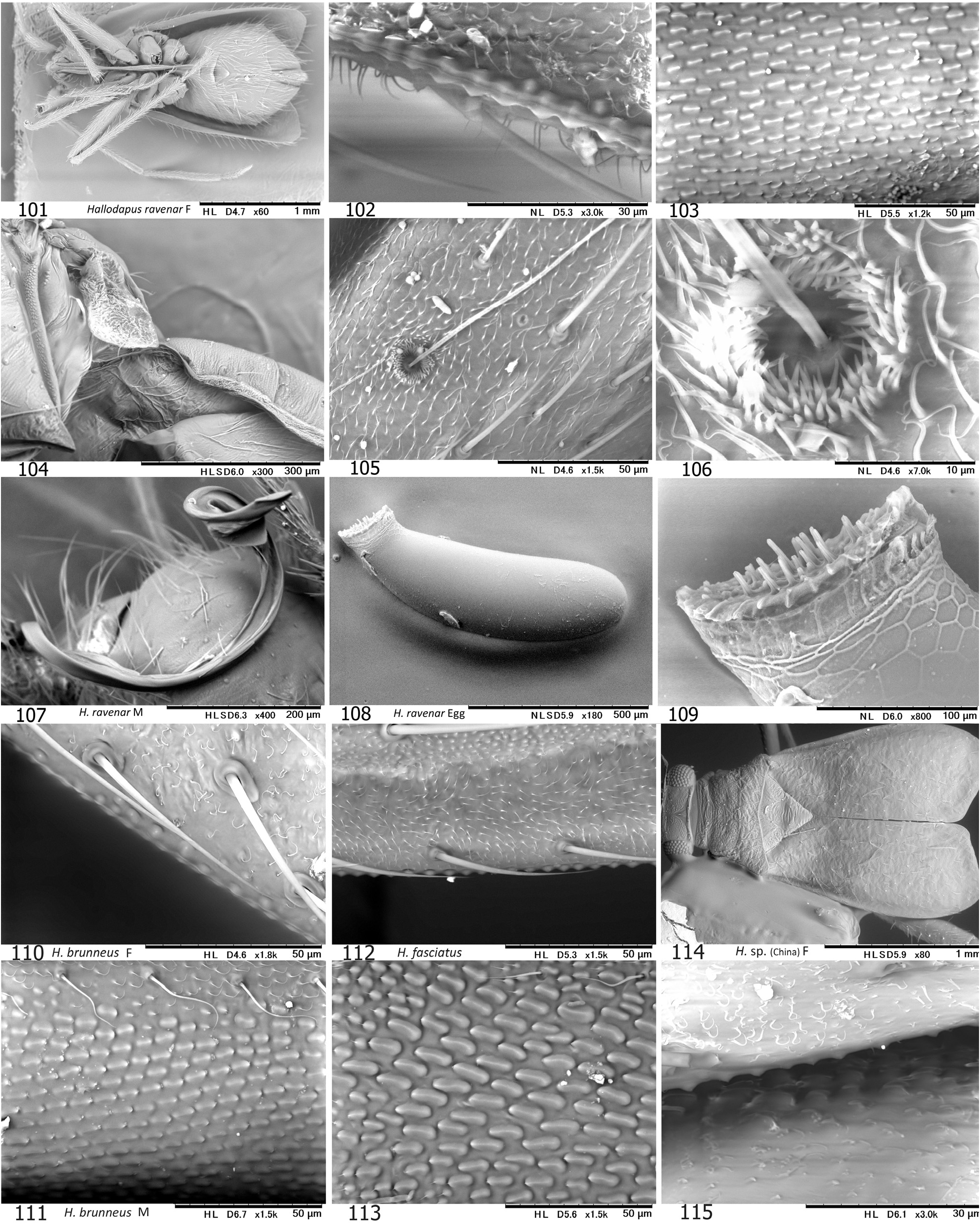
Little has been known about the biology of species grouped into Hallodapini as most available specimens were collected using UV light traps. The immature forms of a few Asian members were found to inhabit and/or propagate on inflorescence and leaves (e.g., Wygomiris kaliyahae , as in Figs 17–18 View Figs 9–18 ), leaves ( Cleotomiroides tobii , Fig. 12 View Figs 9–18 ), tree bark or branches ( Acrorrhinium spp.; see YASUNAGA et al. 2013a, DUWAL et al. 2017) of broadleaf trees ( YASUNAGA et al. 2013a, DUWAL et al. 2017). Thus far as known, most Asian species of Alloeomimella and Hallodapus were confirmed to be epigeic and have the distinct stridulatory device ( Table 1 View Table 1 ). The females of these hallodapines are predominantly brachypterous (forewing coleoptery with rudimentary hindwings, cf. Figs 104 View Figs 101–115 , 216–217 View Figs 201–217 ); the females of Systellonotus species are exceptionally staphylinoidy ( YASUNAGA 2001; WYNIGER 2006). Due to sexual dimorphism of the wings (brachypterous females that cannot take flight), the majority of individuals attracted to UV light are male as a matter of course. The brachypterous males are known in European hallodapines (e.g., Hallodapus montandoni Reuter, 1895 and Omphalonotus quadriguttatus (Kirschbaum, 1856) , see WYNIGER 2006). KMENT & BAŇAŘ (2012) also reported that Hallodapus montandoni is an epigeic species inhabiting xerothermic habitats, mostly on limestone, rarely on sandy ground and possibly associated with ants, especially Myrmica spp.
YASUNAGA & DUWAL (2016) discussed a spine on the genital segment (cf. Figs 30 View Figs 25–31 , 124, 130 View Figs 116–130 , 136 View Figs 131–145 ) that is recognized in some members of Hallodapini ; however, the spine is usually possessed only by males ( Table 1 View Table 1 ). Within Asian hallodapines, presence of the spine in each sex (both P- and O-spines) was confirmed in Clapmarius thailandana and Peniculimiris meniscus . Although the P- and O-spines of these taxa were assumed to stabilize the mating position, the actual functions of these spines are yet to be demonstrated ( YASUNAGA & DUWAL 2016).
On the other hand, the stridulatory device (FWS+MFP) is currently assumed to be synapomorphy for the four Asian genera ( Alloeomimella + Hallodapus +Cleotomiris-+ Wygomiris ) plus two Afrotropical Laemocoris + Trichophthalmocapsus . The well-developed device has hitherto been found only in ground-inhabiting hallodapines. The devices in Cleotomiris miyamotoi ( Figs 66–67 View Figs 56–70 ) and Wygomiris paveli ( Figs 55 View Figs 40–55 , 141–142 View Figs 131–145 ) are obviously reduced and seems to lose its original function. Incidentally, the scent efferent system is noticeably small in two epigeic genera, Alloeomimella and Hallodapus , whereas it is enlarged in Cleotomiris and Wygomiris . The current evidence suggests that the ground-inhabiting hallodapines (in Alloeomimella and Hallodapus ) may utilize sound for intraspecific communication rather than pheromones (from the scent gland). In addition, the parempodia of all the epigeic Alloeomimella and Hallodapus species we examined are setiform; such species frequently slip and walk clumsily on smooth surface (e.g., acrylic or glass Petri dish) but the simplified pretarsal structure possibly enables agile and comfortable movement on the chaotic, messy ground of deep meadows. Nonetheless, much broader morphological and ecological survey treating all hallodapine taxa is required to elucidate the evolution and phylogeny of Hallodapini .






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.