
Megophrys oropedion, Mahony, Stephen, Teeling, Emma C. & Biju, S. D., 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3722.2.2 |
|
publication LSID |
lsid:zoobank.org:pub:0476AFF3-92EC-4649-8FFE-758275EC14E1 |
|
DOI |
https://doi.org/10.5281/zenodo.5696842 |
|
persistent identifier |
https://treatment.plazi.org/id/03CDF20C-FF83-FFA3-7C86-FB9A5CE9F828 |
|
treatment provided by |
Plazi |
|
scientific name |
Megophrys oropedion |
| status |
sp. nov. |
Megophrys oropedion View in CoL sp. nov.
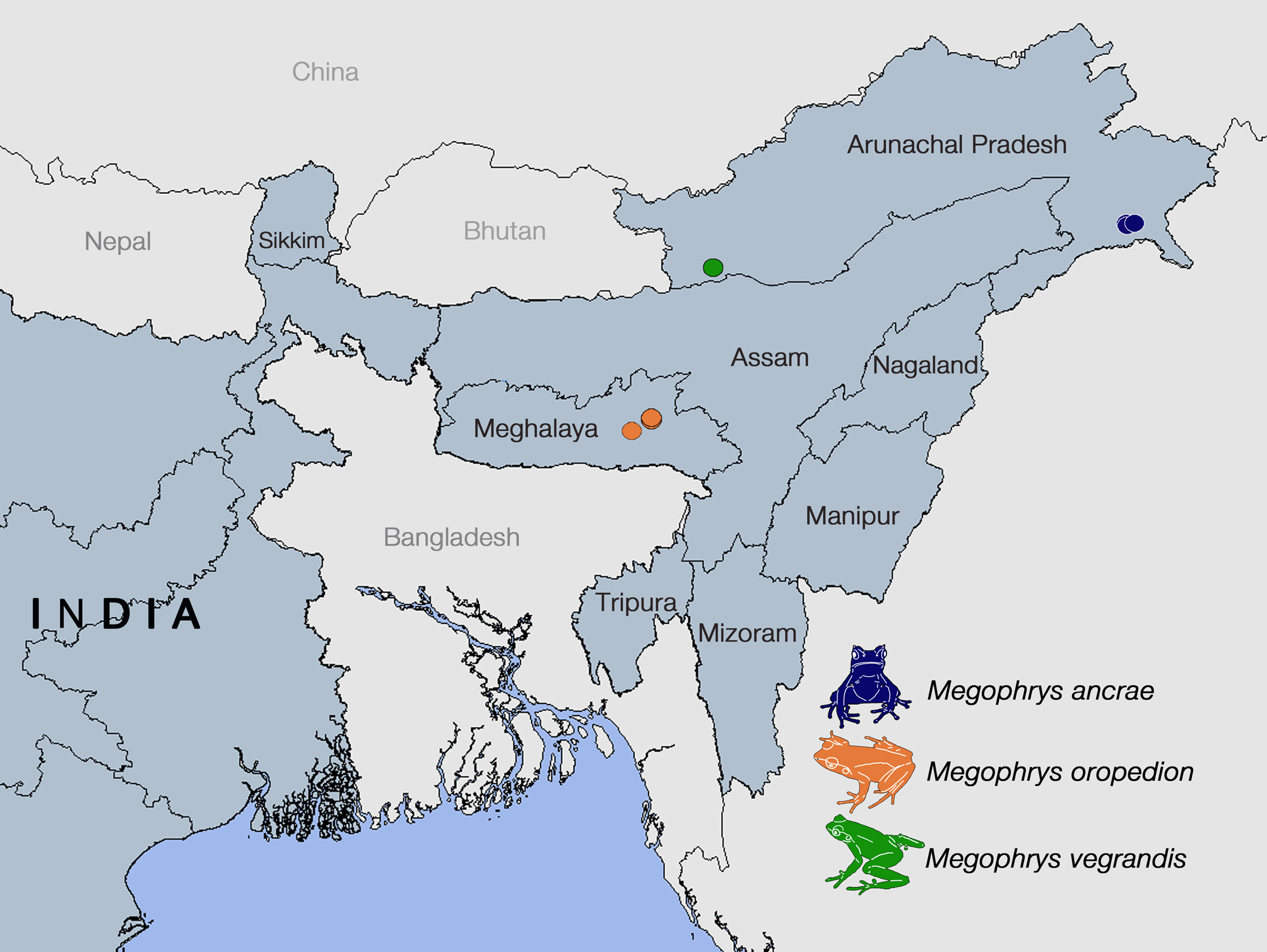
( Figures 3 View FIGURE 3 , 5–7 View FIGURE 5. A – E View FIGURE 6. A – C View FIGURE 7 ; Table 1 View TABLE 1 )
“ Megophrys monticola ”: Pillai and Chanda (1979: 386); Dutta (1997: 37—part; “Shillong Peak”) “ Megophrys parva ”: Chanda (1994: 19, 21—part; “Shillong, Khasi Hills”); Chanda (1995: 459—part; “Shillong”); Dutta (1997: 38—part; “Shillong”); Chanda (2002: 11—part; “Meghalaya”)
“ Xenophrys cf. parva ”: Mahony (2008: 9); Das et al. (2010: 54)
Holotype. Adult male (ZSI A 11601), from Um Risa stream (25°33.644’N 91°52.960’E, 1,520 m asl.), Malki Forest (on Shillong Peak), Shillong, East Khasi Hills district, Meghalaya state, northeast India, collected by Systematics Lab members, 18 May 2009.
Paratypes. Three adult males (ZSI A 11602 View Materials , BNHS 5595, BNHS 5596), collected with holotype; adult female (ZSI A 11603), from Um Risa stream (25°33.327’N 91°53.099’E, 1,600 m asl.), Malki Forest, Shillong, East Khasi Hills district, Meghalaya state, northeast India, collected by Systematics Lab members, 22 May 2009.
Referred specimens. Three adult males (SDB.DU 2009.300–301, SDB.DU 2009.1164) and one adult female (SDB.DU 2009.299), from Mawphlang Sacred Forest (25°26.617’N 91°44.767’E, 1,810 m asl.), Mawphlang, East Khasi Hills district, Meghalaya state, northeast India, collected by Systematics Lab members, 30 May 2009.
Diagnosis. Megophrys oropedion sp. nov. is diagnosable from geographically relevant congeners by the following combination of characters: small-sized, robust species, adult-male SVL 32.8–39.2 mm (N=7), adultfemale SVL 44.1–48.7 mm (N=2); HW≥HL, HW/SVL 33.9–38.5% (N=9); tympanum clearly defined, TYD /ED 53.7–69.0% (N=9); SHL/SVL 41.7–48.4% (N=9), mean male SHL/SVL 45% (N=7), mean female SHL/SVL 42.1% (N=2); vomerine ridges ovoid, with vomerine teeth present; subarticular, inner and outer palmar tubercles and inner metatarsal tubercles absent in life and in preservation; digit tips not expanded relative to digit width; webbing and lateral fringes on toes absent; dorsolateral folds always present, approximately 75 percent of trunk length; distinct “X”-shape marking on dorsum absent; protruding projection posterior to cloaca of males absent; single dominant frequency range of male advertisement call at 3,100–4,250 kHz, with maximum note energy of between 3,364–3,527 kHz (3,388±70 kHz) at 18°C ( Figure 7 View FIGURE 7 ).
Comparisons. Megophrys oropedion sp. nov. differs from the following large-sized species by considerably smaller adult-male size, SVL 32.8–39.2 mm, N=7 (vs. adult-male SVL> 53 mm: M. gigantica , N=6; M. glandulosa , N=10; M. jingdongensis , N=2; M. lekaguli , N=8; M. major , N=8; M. medogensis , N=16; M. omeimontis , N=10; M. robusta , N=4; M. spinata , N=2); from the following medium-sized species by smaller adultmale size, SVL 32.8–39.2 mm, N=7 (vs. Megophrys ancrae sp. nov. male SVL 39.1–45.0 mm, N=8; M. binlingensis SVL 45.1–51.0 mm, N=3; M. megacephala SVL 45.9–53.4 mm, N=7; M. takensis SVL 47.3–53.0 mm, N=3), further from Megophrys ancrae sp. nov. by HW≥HL (vs. HL>HW), digit tips not expanded terminally (vs. digit tips distinctly expanded terminally), small sharp horn-like tubercle absent on the eyelids (vs. present on most individuals), further from M. binlingensis by lateral fringes on toes absent (vs. present), further from M. megacephala by considerably narrower head HW/SVL 33.9–38.5%, N=9 (vs. HW/SVL 40.2–45.1%, N=9), and further from M. takensis by vomerine ridges ovoid (vs. elongated stalk-like projections); from the following smallsized species by larger adult size, male SVL 32.8–39.2 mm, N=7, female SVL 44.1–48.7 mm, N=2 (vs. M. wuliangshanensis male SVL 27.3–31.6 mm, N=10, female SVL 41.0– 41.5 mm, N=2; M. zunhebotoensis male SVL 30.0 mm, N=1, female SVL 39.0 mm, N=1), further from M. wuliangshanensis by larger tympanum on males, mean TYD /ED 60.4%, N=7 (vs. mean male TYD /ED 48.8%, N=10), considerably shorter mean shank length, mean male SHL/SVL 45%, N=7, mean female SHL/SVL 42.1%, N=2 (vs. mean male SHL/SVL 50.5%, N=10, mean female SHL/SVL 50.4%, N=2), and further from M. zunhebotoensis by male SHL/SVL 43.4–48.4%, N=7, female SHL/SVL 41.7–42.6%, N=2 (vs. male SHL/SVL 50%, N=1, female SHL/SVL 46.1%, N=1); from M. nankiangensis and M. shapingensis by presence of clearly defined tympanum (vs. tympanum concealed by supratympanic fold); from M. binchuanensis and M. wushanensis by subarticular tubercles absent (vs. present), and lateral fringes on toes absent (vs. present); from M. minor by HW≥HL (vs. HL>HW), vomerine teeth present (vs. absent), differences in male advertisement call characters, note duration 30–55 ms (44.6±7.33 ms, N=32) and a single primary frequency range at 18°C (vs. note duration 74.6–110.1 ms [90.8±12.6 ms, N=14], and a second subdominant frequency pulse at 6,800–7,600 kHz at 14°C); from M. palpebralespinosa by webbing between toes absent (vs. approx. half digit length); from M. daweimontis by inner and outer palmar tubercles and inner metatarsal tubercles absent in life and in preservation (vs. distinct and red in colour), SHL/SVL 41.7–48.4%, N=9 (vs. SHL/SVL 53.7% on holotype); from M. wawuensis and M. zhangi by distinct “X”-shape marking on dorsum absent (vs. present) and dorsolateral folds distinct (vs. indistinct), further from M. wawuensis by larger tympanum, TYD /ED 53.7–69.0%, N=9 (vs. TYD /ED ca. 33%), and further from M. zhangi by lateral fringes on toes absent (vs. present); from M. pachyproctus by protruding projection posterior to cloaca of male absent (vs. present); from M. parva by larger adult female size, SVL 44.1–48.7 mm, N=2 (vs. female SVL 41.1–41.4 mm, N=2), larger tympanum TYD /ED 53.7–69.0%, N=9 (vs. TYD /ED 40.0–48.9%, N=4), and digit tips not expanded relative to digit width (vs. digit tips noticeably expanded relative to digit width); from M. serchhipii by dorsolateral folds always present, approximately 75 percent of trunk length (vs. absent).
Holotype description (measurements in mm). Mature male (SVL 32.8) ( Figure 5 View FIGURE 5. A – E , 6C View FIGURE 6. A – C ). Head small (HW 12.0, HL 12.0, IFE 6.0, IBE 10.1), as wide as long; snout rounded in dorsal view, obtusely protruding in lateral view, without rostral appendage ( Figure 5C View FIGURE 5. A – E ); loreal region vertical and concave; canthus rostralis angular; dorsal region of snout slightly concave; eye (EL 4.4) longer than maximum tympanum diameter ( TYD 2.7), and subequal to snout (SL 4.5); eye-tympanum distance (TYE 1.8) shorter than maximum tympanum diameter; tympanum slightly oval, vertically orientated, not concealed by supratympanic ridge ( Figure 5C View FIGURE 5. A – E ); pupil in life oval, horizontally orientated when dilated and vertically when constricted; nostril orientated laterally, situated mid-way between eye and snout (EN 2.2, SN 2.3); internarial distance (IN 3.8) equal to eyelid width (UEW 3.8), and wider than the narrowest point between upper eyelids (IUE 3.4); pineal ocellus not visible externally; vomerine ridges moderately raised, ovoid, positioned at level posterior to choanae, separated from each other by distance equal to distance from choanae; vomerine teeth small; tongue moderately large, weakly notched posteriorly, with no medial lingual process.
Forelimbs moderately short and thin, forearm (FAL 7.6) slightly enlarged relative to upper forelimb, shorter than hand (HAL 10.1); fingers short and rounded without lateral fringes ( Figure 5D View FIGURE 5. A – E ), finger length formula I=II=IV<III (FIL 3.9, FIIL 4.0, FIIIL 6.3, FIVL 4.0); interdigital webbing, thenar, palmar, subarticular and supernumerary tubercles absent; finger tips not expanded, rounded with small oval pads; terminal grooves absent. Hindlimbs relatively short and thin; shanks overlap when thighs are held at right angle to body, thigh (TL 13.9) shorter than shank (SHL 14.4), and foot (FOL 14.5); toes long and rounded without lateral fringes ( Figure 5E View FIGURE 5. A – E ); relative toe lengths I<II<V<III<IV; toe tips not dilated, but with small terminal pads; terminal groves absent; outer metatarsal tubercle, subarticular and supernumerary tubercles and webbing absent; inner metatarsal tubercle indistinct; ridge of callous tissue absent on the ventral surface of digits.
Skin of dorsal surfaces of body, limbs, and dorsal and lateral surfaces of head weakly granular; tympanum smooth, its borders slightly raised; short ridge present on outer edge of upper eyelid ( Figure 5C View FIGURE 5. A – E ); supratympanic fold extends from orbit and curves around upper border of tympanum, terminating above insertion of forelimb ( Figure 5C View FIGURE 5. A – E ), its posterior half more than 50 percent wider than anterior half; flanks with large scattered tubercles; thin dorsolateral folds extend from posterior to supratympanic fold to approximately three quarters distance to groin; weak, “V”-shaped parietoscapular ridge present, its two sides extending posteriorly from above tympanum and meeting medially beyond level of axilla; second inverted partial “V”-shaped ridge present on mid-dorsum, joining laterally with dorsolateral fold on right side only ( Figure 5A View FIGURE 5. A – E ); small tubercles arranged into distinct transverse rows on dorsal surface of thighs, shanks and forearms. Gular region, chest and ventral surfaces of limbs smooth; abdomen weakly granular; pectoral glands moderately large and raised slightly, positioned on level with axilla ( Figure 5B View FIGURE 5. A – E ); femoral glands moderately large, slightly raised, positioned sub-equally distant from knee and cloaca. Conspicuous brown skin asperities form a narrow band circummarginally on lower jaw, densely scattered at posterior end of jaw and on tympanic region (except tympanum), along upper jaw, loreal region and snout, and on supratympanic and dorsal folds; sparse on head, dorsal surface of snout and anterior half of dorsum, dorsal surfaces of forelimbs and hindlimbs, posterior half of upper eyelids, and upper flanks; dense on the posterior half of dorsum, and surrounding cloaca, extending ventrally onto posterior part of abdomen; few asperities extending from axilla to adjacent pectoral region including the pectoral glands; absent from remaining surfaces.
Colour in preservative ( Figure 5 View FIGURE 5. A – E ): Entire dorsal and lateral surfaces of head, body, forelimbs and hindlimbs light brown; solid darker brown triangular marking between eyes, and thin dark brown stripe extending from tip of snout posterodorsally to centre of snout; supratympanic folds whitish-cream; front of snout with dark brown blotches; wide vertical dark brown bar below eyes and dark brown blotches covers tympanum; two dark brown blotches on anterior lateral surface of forearm, and dorsal surface of hands and feet with dark brown spots and speckles; feint dark brown transverse bands present on dorsal surfaces of hindlimbs. Gular region, chest, abdomen and ventral surface of limbs pale greyish-brown; feint, dark mottling on limbs; some small white spots along margin of lower mandible and distinct dark brown patch on either side of posterior ends of mandibles extending to insertion of forelimbs; ventral surfaces of tarsus and feet dark grey brown; area surrounding vent and posterior surfaces of thighs dark brown; ventral surface of hands grey-brown; pectoral and femoral glands white. Colour in life ( Figure 6C View FIGURE 6. A – C ): As described above, except that dorsal colour was a richer mid-brown with tips of dorsal tubercles and large granules orange; inner surface of thighs and groin with some orange colouration. Iris colour metallic brown.
Variation. See Table 1 View TABLE 1 for morphometric characters of seven adult males and two adult females. Finger length formula varies considerably in this species with the following combinations found in the series examined: I≤II≤IV<III (N=6), II<IV<I<III (N=1), and I<IV<II<III (N=2). ZSI A 11603 has considerably smaller, ovoid vomerine ridges, and SDB.DU 2009.1164 possesses only one small, ovoid vomerine ridge on the right side, with the left vomerine ridge absent. Vomerine ridges in the remaining specimens like in the holotype. Dorsolateral folds are present on all examined specimens and vary in length from approximately 75–100 percent of the trunk length. Tympanum is completely exposed on five specimens but on four the supratympanic fold covers the dorsal-most border, and the tympanum shape varies from circular to distinctly vertically oval. Asperities density and distribution for all specimens are similar to the conditions described for the holotype with the following exceptions: BNHS 5595 differs from the holotype by having only a single asperity on each pectoral and femoral gland; ZSI A 11603 has sparse, white asperities on supratympanic and dorsal folds, at the rear of the jaw, on posterior portions of eyelids, and dorsal surfaces of forelimbs and hindlimbs, dense asperities on the posterior half of the dorsum and the surroundings of the cloaca, and two sparse patches on the chest adjacent the axilla, but absent from other surfaces; SDB.DU 2009.300 differs from the holotype by having black and white asperities on the lower jaw, and axillary asperities absent; SDB.DU 2009.301 differs from the holotype by having a broad band of asperities along the margin of the lower jaw. Most specimens match the holotype by possessing a short raised dermal ridge on the upper eyelid except BNHS 5595 and SDB.DU 2009.301 which have a small blunt tubercle on each eyelid replacing the ridge, and SDB.DU 2009.299 which has a ridge on one side and a tubercle on the other. The parietoscapular ridge on five of the nine specimens does not join medially to form a “V” shape, but appears as “\ /”.
Colouration and markings of the type series and referred specimens generally agree with the holotype with the following exceptions: all other specimens have a distinct longitudinally elongated brown patch on the central gular region extending onto the anterior part of the chest; some individuals have additional large dark brown blotches along the margin of the lower jaw and dense, darker brown blotches on the dorsum (e.g., SDB.DU 2009.301, ZSI A 11602 View Materials , BNHS 5595, BNHS 5596); dark bands on the dorsal surfaces of the hindlimbs and stripe on the dorsal portion of the snout indistinct on SDB.DU 2009.1064; the entire gular, chest and ventral abdominal region of SDB.DU 2009.300–301 and BNHS 5595 are covered with feint to dark brown blotches and mottling; flank tubercles in general are lighter than surrounding areas. Many individuals of this species that were photographed but not collected from the localities listed in the distribution section varied in general dorsal colouration from dark brown to bright orange; the inner surfaces of the thighs and groin of males and females were orange to reddishorange; the aforementioned feint brown blotches on the ventral surface in preservative are often orange in life.
Secondary sexual characters. Males have slightly raised nuptial pads covered with black micro-granules, covering most of the dorsal surface of the base of finger I, narrowing distally and extending to the base of the distal phalange on the inner dorsal side; nuptial pad medium sized and oval shaped on finger II, on the inner dorsal side of the base of the digit extending to the mid-proximal phalange; external vocal sac distinct as loose skin on most specimens, and forms a large single subgular sac that extends onto the anterior chest when fully inflated; internal vocal slit present on the floor of the mouth, near the rear of the lower mandible on both sides; forearm slightly to moderately enlarged relative to upper forelimb; protruding fleshy projection posterior to cloaca absent. Females lack nuptial pads, external vocal sac, internal vocal slits and enlarged forearms—ova are unpigmented.
Etymology. The specific epithet “ oropedion ” is Greek meaning “plateau” or “tableland”, as all known collection localities for the species are situated on the upper reaches of the Shillong Plateau. We treat the specific epithet as a noun in apposition to the generic name.
Distribution. This species is confirmed here from Malki Forest (1,520–1,600 m asl.), Shillong, and Mawphlang Sacred Forest (1,810 m asl.), Mawphlang village, both in the East Khasi Hills district of Meghalaya state, northeast India ( Figure 3 View FIGURE 3 ). It has also been observed, but not collected from North East Hill University (NEHU) (25°36.767’N 91°45.050’E, 1,410 m asl.), on the outskirts of Shillong city (SM pers. obs.). This species is likely to be found in other forest fragments in the East and West Khasi Hills districts that cover the upper reaches of the Shillong Plateau above 1,400 m asl.
Habitat and natural history. The forest type around Shillong and Mawphlang is categorized as subtropical wet hill forest (Champion & Seth 1968). The forest at the type locality is dominated by pine ( Pinus kesiya Royle & Gordon ) plantation, with a considerable amount of broadleaf trees lining the stream banks (SM, pers. obs.) ( Figure 4B View FIGURE 4. A – D ), while the forest at Mawphlang is dominated by broadleaf trees ( Figure 4C View FIGURE 4. A – D ). The collection streams at these localities were approximately 1–3 m wide and 10–40 cm deep, of moderate flow on a slight incline, primarily rocky with intermittent gravel bed, and with quiet pools that accumulate detritus, branches and leaf litter (for further details on habitat at Mawphlang see Mahony [2008]). The species was only found along portions of the streams that experience low levels of human disturbance. One female was collected on a trail ca. 200 m from the nearest stream, and another collected from a trail nearby a stream. Males were collected from the lower branches of bushy vegetation at ca. 20 cm above ground level, within 2 m from the edge of streams, or from rocks on the stream banks. One uncollected male was located inactive during day time in a fallen hollow branch lying over a stream (1 m above stream level). Metamorphosing juveniles were found during May and June at Shillong and Mawphlang. Males began calling vigorously from dusk until late into the evening (18:00 until at least 23:30 h), regardless of precipitation level, but more intensely during light rain and/or when cloud cover envelops the forest, when regular calling began as early as 16:00 h, but was reduced during heavy precipitation. Calling was observed during visits between 20th May to 15th September. Sporadic daytime calling was heard on both overcast and clear days at Shillong and Mawphlang, and it consisted of one to two call repetitions followed by extended silence, usually from unobserved calling sites within dense bushes. Calling males distanced themselves from others by a minimum of 1 m of stream bank and distinctive territorial calls were emitted when males were played back their call at close quarters, as noted in other species (Mahony & Reza 2008).
The advertisement call of Megophrys oropedion sp. nov. was recorded from two individual males (not collected) from the banks of the Um Risa stream within meters of the collection locality of the holotype. These individuals were photographed in-situ and released, and match the type series in all general external morphological characters. The call consists of a single pulsed note of 30–55 ms duration (44.6±7.33 ms, N=32), containing 6–10 pulses per note (7.7±0.97, N=32). Pulse intervals gradually decrease from the beginning to the end of the note. Amplitude modulation within notes is apparent, beginning with moderately high energy pulses, increasing slightly to a maximum by approximately mid note, and subsequently decreasing rapidly towards the end of each note. Calls are repeated in series at a rate of 3–4 per second, with an irregular call interval of 108–415 ms duration (276±67 ms, N=32). Duration of call series was not recorded, but calls were continuously emitted throughout the longest recording made of 24 s. Calls had a single dominant frequency range of 3,100–4,250 kHz with the maximum note energy varying from 3,400–3,500 kHz (3,388±70 kHz, N=27).
Out of a total of 18 metamorph to adult frogs photographed, two examples with eye deformities were observed (not collected), one from Mawphlang and the other from the lower Malki forest. In both cases, one eye was smaller than the other, with the pupil off-center and partially obscured by the upper eyelid. It was not tested at the time of observation whether the deformed eyes were functional. At both of these localities, the breeding streams are intensively used as clothes-washing areas and in such sections of the streams the rocks are covered with a grey slime from excessive detergent use and calling males are absent (see Mahony 2008 for further discussion). Whether detergent use in these streams is responsible for causing the malformations, or whether it is a result of natural parasite/viral infection or trauma should be further investigated.
Remarks. Pillai and Chanda (1979) report a single specimen in the ZSI/ERS which they refer to as “ Megophrys monticola Kuhl. & V. Hass., 1822 [b]” (= Megophrys montana Kuhl & Van Hasselt, 1822 [a]), collected from Shillong Peak—the type locality of Megophrys oropedion sp. nov. Immediately obvious is their confusion with the name Xenophrys monticola Günther, 1864 (currently a synonym of Megophrys parva ), the name which they presumably intended to refer to based on later corrections of the identification of this specimen to M. parva (Chanda 1994, 1995, 2002). Pillai and Chanda (1979) provide a basic description of this specimen which corresponds with Megophrys oropedion sp. nov., with the exception that they identify the specimen as a male with SVL of 48 mm (vs. male SVL 32.8–39.2 mm, N=7, reported herein). This measurement however corresponds perfectly with females of this species, thus the sex of this specimen should be re-examined. Dutta (1997) lists both Megophrys monticola Kuhl. & V. Hass., 1822 (= Megophrys montana Kuhl & Van Hasselt, 1822 [a]) and Megophrys parva from Shillong presumably based on the aforementioned papers (Pillai & Chanda 1979; Chanda 1994), as both references are included in the bibliography section. Considering Megophrys oropedion sp. nov. was consistently the only species found on Shillong Peak during numerous visits by the Systematics Lab members between 2006–2011, we regard the specimen of Pillai and Chanda (1979) as this species. Megophrys montana Kuhl & Van Hasselt, 1822 [a] is otherwise known from the Indonesian island of Java (Inger 1954), and is therefore not a member of the Indian amphibian fauna despite being listed as such several times in the past (e.g., Pillai & Chanda 1979; Dutta 1997; Das & Dutta 1998). Mahony (2008) reported Megophrys oropedion sp. nov. (as Xenophrys cf. parva ) from Mawphlang, which was later cited by Das et al. (2010).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.