
Haplodidymos rubroculatus, Hochberg, Rick & Cannon, Lester R. G., 2002
|
publication ID |
https://doi.org/ 10.5281/zenodo.156078 |
|
DOI |
https://doi.org/10.5281/zenodo.6279085 |
|
persistent identifier |
https://treatment.plazi.org/id/03CE878B-FF80-5D28-E20F-F9D521377D6D |
|
treatment provided by |
Plazi |
|
scientific name |
Haplodidymos rubroculatus |
| status |
sp. nov. |
Haplodidymos rubroculatus View in CoL sp. nov.
Material. Eleven animals studied alive, six mounted, and three serially sectioned.
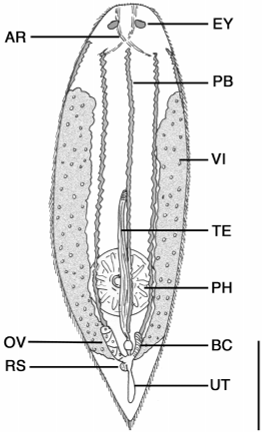
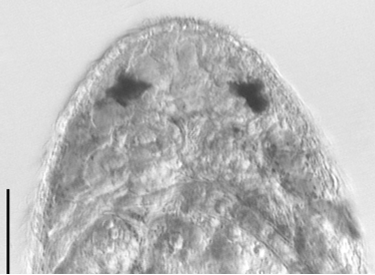
Description. Animals are up to 720 µm long with a rounded head and tapering caudal end that forms a small tail ( Fig. 7 View FIGURE 7 ). The worms are highly flexible and often contract into a spherical shape when disturbed. Worms are mostly transparent along their length, though several longitudinal bands of redbrown pigment are present around the body. During full body contraction, the pigment gets dispersed around various internal organs. Epidermis consists of cuboidal cells, 8 µm high, with cilia to 10 µm long. The head contains a large (24 µm long) pair of red eyes ( Fig. 8 View FIGURE 8 ). Individual pigment granules were not observed. The eyes were not present after fixation in de Faure’s fluid. In section, the region around and including the eyes stains intensely with eosin. Immediately posterior of the eyes is the cerebral ganglion; cerebral cells stain intensely with aniline blue. Paired tracts of adenal rhabdites extend for up to 136 µm from the tip of the head past the eyes. The tracts are spaced widely at the tip of the head and connect posterior to the eyes with lateral branches extending toward the body margin.
The digestive tract consists of a saccate gut and pharynx rosulatus. The gut often contain numerous rotifer trophi and occasionally full rotifers – species of Philodina and various ploimate rotifers. Gut cells are eosinophilic. The vertically oriented pharynx rosulatus is present in the posterior onethird of the body and up to 128 µm diameter. Protonephridia pores were not evident in wholemount or section material. Caudal glands stained with the basic dye aniline blue indicative of acid mucosubstances.
The female reproductive system consists of paired vitellaria, a solitary ovary, receptaculum seminis, bursa copulatrix, uterus, genital atrium and common gonopore ( Fig. 9 View FIGURE 9 ). The paired vitellaria are papillose and begin around 25% body length and extend to ca. 85% body length. Vitelline ducts may form a single duct before opening close to the base of the oviduct or perhaps the lateral wall of the genital atrium. The solitary ovary is variable in position but often located medially, extending dorsal or sometimes ventral to the gut. The ovary opens at its posterior tip into a thinwalled oviduct that leads to the genital atrium. The pouchlike genital atrium has a single midventral gonopore present at ca. 90% body length. A circular receptaculum seminis, ca. 3040 µm diameter, opens into the left wall of the genital atrium through a short duct, and an elongate bursa copulatrix, ca. 5064 µm long, opens into the right wall. The bursa is highly muscular with strong circular muscles and weaker longitudinal bands. Both organs were filled with sperm in several specimens. A thin sacshaped uterus connects to the posterior wall of the genital atrium and contains a single egg. Eggs are bright orange, devoid of sculpture, and up to 46 µm long by 34 µm wide.
The male reproductive system consists of a solitary testis, male copulatory organ, genital atrium and common gonopore ( Figs. 7 View FIGURE 7 , 9 View FIGURE 9 ). The single testis is located ventral to the gut along the body midline. It is fingerlike in appearance and up to 120 µm long. The anteriormost portion of the testis is highly compact while free sperm are present posteriorly along most of its length. Distally, the testis curves dorsally and opens into a short vas deferens before entering the anterior border of a compact male copulatory organ. The copulatory organ is dorsally located and has two distinct portions, an anterior seminal vesicle and posterior copulatory bulb. The seminal vesicle is a thinwalled oblong organ, ca. 16 µm wide, and often filled with sperm. The seminal vesicle sits atop a glandulomuscular organ containing a weakly sclerotic stylet. Prostatic glands were not observed outside of the organ, nor was there a defined glandular mass within the organ. Instead, finelygranular refractive spheres (glandular?) were scattered throughout the organ and around the ejaculatory duct containing the stylet. The stylet is weakly curved and to 31 µm long. Proximally, the stylet opens to 4.2 µm wide, then narrows to 2.5 µm, and expands to 5.4 µm at the distal opening.
Remarks. Several characters link this species with other members of Typhloplanidae Graff, 1905 : the pharynx rosulatus, single ovary with separate follicular vitellaria, and single gonopore. There are currently eight subfamilies within the Typhloplanidae Graff, 1905 , generally all defined by the orientation and position of the pharynx, location of excretory pores, and position of testes relative to vitellaria ( Cannon 1986). The taxonomic position of Haplodidymos can be defined by these and other characters including the presence of eyes, uterus, and the structure of the stylet.
The ventral orientation of the pharynx rosulatus in Haplodidymos is characteristic of most species of Typhloplanidae with the exception of species of Phaenocorinae and Opistominae , where the pharynx is anteriorly and posteriorly directed, respectively. The location of the pharynx in the posterior onethird of the body is characteristic of species of Olisthanellinae , Protoplanellinae and Typhloplaninae .
The location of excretory pores was difficult to identify conclusively in the specimens examined. Based on limited observations, protonephridial ducts were never observed to enter the region of the mouth (e.g., Mesostominae , Typloplaninae).
Eyespots are common among typhloplanids with the exception of species of Typhloplana and Typhloplanella . The eyespots of Haplodidymos appear to be unique among Typhloplanidae . Instead of consisting of numerous pigment granules forming a distinct cup, the eyes consist of large patches of red pigment without distinct granules. Closer inspection is necessary before speculating on their precise structure.
The general structure of the reproductive system is similar to other typhloplanids with three notable deviations: number and position of testes, structure of the male copulatory apparatus, and the presence of a uterus. The presence of a solitary testis lying along the body midline in Haplodidymos is an unusual and unique condition among species of Typhloplanidae . Anatomically and histologically, the testis is a long thin organ, with a compact, proximal germinal zone, and distal, slightly dilated zone of free sperm. The latter zone takes up most of the length of the organ and may in fact represent a highly dilated vas deferens. Only at its most distal tip does the testis form a narrower zone reminiscent of the thin vas deferens of other typhloplanids. Sperm congregate at the distal end of the testis where it connects with the anterior wall of the copulatory organ and forms a vesicula seminalis. The ventral position of the testis relative to the vitellaria suggests close ties with the Protoplanellinae , Rhynchomesostominae and Typhloplaninae .
The second deviation in the reproductive system is in the structure of the copulatory apparatus. According to Jondelius and Thollesson (1993), a cirrus is characteristic of the ground pattern of the family. However, Haplodidymos possesses a simple stylet, as do species of the marine genera Brinkmanniella , Haloplanella , Pratoplana , and Thalassoplanella among others. The stylet of Haplodidymos is weakly developed relative to these other species. The presence of a stylet may be indicative of close phylogenetic ties. The third deviation from the reproductive ground pattern of the Typhloplanidae is in the presence of a uterus ( Jondelius & Thollesson 1993). Egg maturation probably occurs in the parenchyma of most typhloplanids, while in Haplodidymos and several species of Chorizogynopora ( Kolasa 1980) , Olisthanella and Phaenocora ( Luther 1963) , a posterior uterus is present. Other species such as Gullmariella vivipara Luther, 1948 possess a superior genital atrium for egg development. However, apart from their respective positions, the difference between the uterus and superior genital atrium is ambiguous according to the diagrams of Luther (1948) and Kolasa (1991). In Luther’s scheme, the superior genital atrium is a lone, dorsal expansion of the genital atrium (inferior), but according to Kolasa (1991), the superior genital atrium receives the ducts of other reproductive organs and the uterus functions in egg maturation.
Based on the above characters, Haplodidymos has close taxonomic ties to Protoplanellinae . The precise position of excretory pores remains to be determined, but their absence from the mouth region excludes this genus from Typhloplaninae .

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |