
Algerophilus hispanicus ( Meinert, 1870 )
|
publication ID |
https://doi.org/ 10.5281/zenodo.280399 |
|
DOI |
https://doi.org/10.5281/zenodo.6166526 |
|
persistent identifier |
https://treatment.plazi.org/id/03CE9A2B-195E-AE58-FF6E-551C2626A41A |
|
treatment provided by |
Plazi |
|
scientific name |
Algerophilus hispanicus ( Meinert, 1870 ) |
| status |
|
Algerophilus hispanicus ( Meinert, 1870)
Geophilus hispanicus: Meinert 1870 (p. 70 original description); Daday 1889 (p. 145); Blanchard 1898 (p. 458); Brolemann 1924 (p. 195); Attems 1927 (p. 238), 1929 (p. 182 diagnosis); Jeekel 2003 (p. 93).
Geophilus (Geophilus) hispanicus: Verhoeff 1896 (p. 88 key); Attems 1903 (p. 169, 219 key, 224).
Algerophilus hispanicus: Brolemann 1925 (p. 250 new record and diagnosis), 1931 (p. 311), 1932 (p. 41 key), 1947 (p. 174 new records).
Nesogeophilus Mateui : Machado 1953 (p. 79 original description, figs 1–2). Syn. n.
Nesogeophilus hispanicus: Demange 1963 (p. 88).
Tuoba hispanica: Sammler et al. 2006 View in CoL (p. 302 new records)
Type material. Syntypes: 17 specimens, including 11 Ƥ, 4 33 and 2 other specimens of unknown sex, all from “Sevilla, Granada ”, Spain; in the Zoological Museum, Natural History Museum of Denmark, Univ. Copenhagen.
Notes on type material. A total of only 11 specimens, including 7 Ƥ and 4 33, was given by Meinert (1870), but no evidence exists against the fact that all 17 specimens comprise the original material upon which Meinert described the species, so that we are confident in recognising all of them as syntypes. Two different specimens have been labeled as “ lectotype ” by R. E. Crabill (dated 21.IV.1960) and D. Foddai (dated 9.X.2001), but – at the best of our knowledge – neither student ever published the designation. Contrary to the intent of both previous students, we refrain from selecting a lectotype for G. hispanicus because such an action is not necessary to promote nomenclatural stability (International Code of Zoological Nomenclature: Recommendation 74G).
Synonyms. Nesogeophilus mateui Machado, 1953 syn.n. Type material: holotype 3, from Benaojan, near Malaga, Spain.
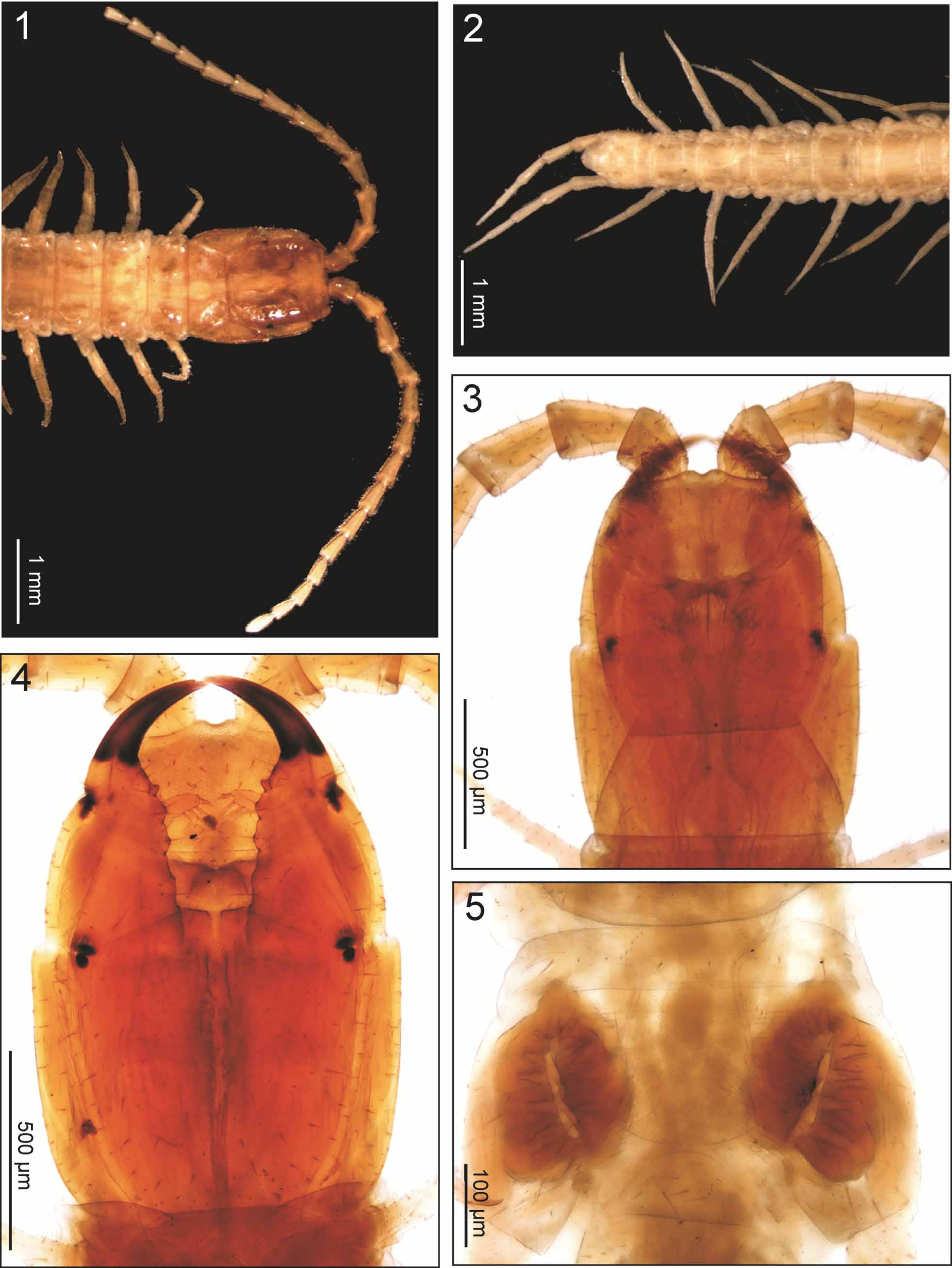
Description of an adult female. One of the syntypes, from Sevilla, Granada, 51 mm long, with 61 leg pairs ( Figs 1, 2 View FIGURES 1 – 5 ; 6–9; 11).
Colour (in ethanol; Figs 1, 2 View FIGURES 1 – 5 ). Almost uniformly light orange-brown, with forcipular condyles and distal part of the forcipular tarsungula dark brown.
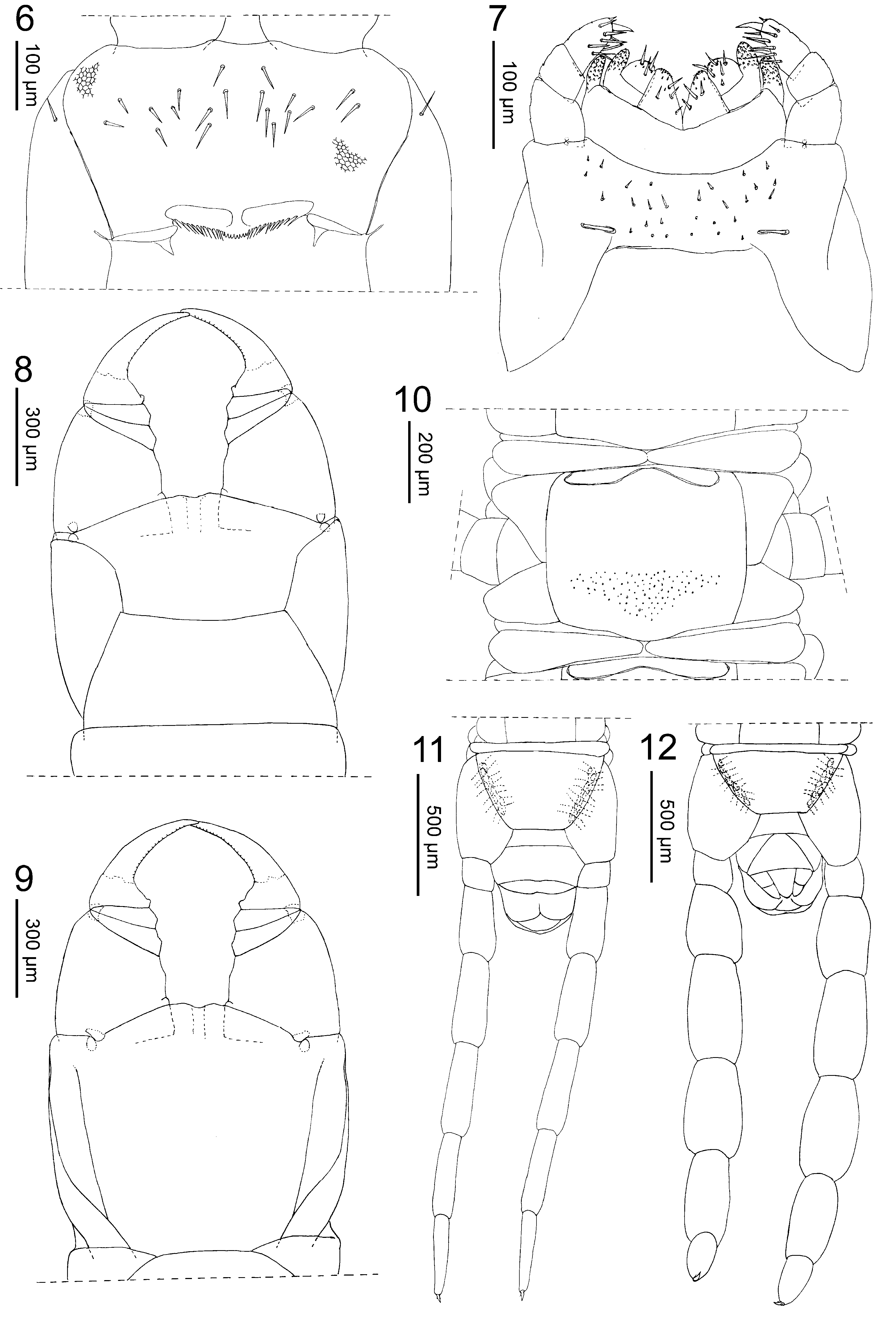
Cephalic capsule. Cephalic plate subrectangular, ca 1.2 times as long as wide, 0.9 times as wide as the forcipular tergite, the anterior margin slightly angulated with a medial notch, the lateral margins distinctly convex, the posterior margin straight; transverse suture absent; setae up to ca 100 μm long. Clypeus ( Fig. 6 View FIGURES 6 – 12 ) uniformly areolate, without clypeal areas and without plagulae; lateral margins complete; a total of 19 setae, of which eight approximately in four medial pairs, the others lateral to them in the anterior half of the clypeus. Pleurites uniformly areolate, each with a single seta in the anterior part. Labrum composed of an intermediate part that is continuous with the clypeus and bears a row of 9 sclerotised short tubercles, and lateral parts that are marginated anteriorly by a complete sulcus and bear a row of 12 hyaline bristles each.
Antennae. Slender, ca 4.2 times as long as the head. Intermediate articles up to ca 2.0 times as long as wide. Article XIV ca 2.7 times as long as wide, ca 1.2 times as long as article XIII and as wide as the latter. Setae up to ca 110 μm long, becoming gradually denser and shorter towards the intermediate articles, both ventrally and dorsally. Apical sensilla ca 10 μm long, spear-like, without projections, only gently narrowing at about the mid-length. Clublike sensilla ca 12 μm long, on article XIV only, grouped on the distal parts of both the internal and external sides. Three longitudinal rows of 1–4 propioceptive spine-like sensilla at the bases of articles; only a single, dorsal sensillum on articles VI, X and XIV. Groups of 1–3 sensilla, similar to the apical ones, ca 8 μm long, on both dorsoexternal and ventro-internal position close to the distal margin of articles V, IX and XIII.
Mandibles. A single pectinate lamella on each mandible.
First maxillae ( Fig. 7 View FIGURES 6 – 12 ). Coxosternite entire, without mid-longitudinal sulcus, without setae. Coxal projection sub-triangular, longer than wide, bearing a few setae and some spine-like sensilla, the tip covered with fine scales. Telopodite composed of two articles, the basal one without setae, the distal one with some setae and some spinelike sensilla, the tip covered with fine scales. Coxosternal and telopodital lappets present, long.
Second maxillae ( Fig. 7 View FIGURES 6 – 12 ). Coxosternite entire, the intermediate part uniformly sclerotised as the remaining parts; anterior margin widely concave, postero-lateral margins pointed; no sclerotised ridges; many short setae on the intermediate part and close to the anterior margin; metameric pores featuring as transversal slits, close to the postero-medial margin. Telopodite composed of three articles, narrowing towards the tip; pretarsus in the shape of a simple claw, subconic, its distal half slightly bent.
Forcipular segment ( Figs 8–9 View FIGURES 6 – 12 ). Tergite subtrapezoid, the lateral margins almost straight and evidently converging anteriorly, ca 2.2 times as wide as long, posteriorly almost as wide as the subsequent metatergite. Pleurites without scapula; lateral margins almost straight and slightly converging posteriorly. Exposed part of the coxosternite ca 1.2 times as wide as long; anterior margin slightly projecting anteriorly with an intermediate shallow concavity, without denticles; one pair of small setae on the dorsal surface close to the anterior margin; coxopleural sutures complete, subparallel on the anterior half, running along the margin on the anterior fourth; chitin-lines distinct, however not reaching the condyles but pointing lateral to these. Basal distance between the forcipules ca 0.2 of the maximum width of the coxosternite. Forcipular trochanteroprefemur approximately 1.1 times as long as wide, the internal side much shorter than the external side, with two shallow mesal bulges. Forcipular intermediate articles distinct, each with a pointed shallow bulge only. Tarsungulum 2.4 times as long as wide, uniformly concave along both internal and external margin; a basal subconic tubercle; internal margin crenulated with ca 20 small irregularly spaced notches. Poison calyx slightly elongate, lodged inside the intermediate articles.
Leg-bearing segments. Metatergite 1 only slightly wider than the subsequent ones, lateral margins converging posteriorly, without pretergite. No paratergites. Metasternites slightly wider than long on the anterior part of the trunk, longer than wide on the posterior part; a distinct bilobed carpophagus pit on some anterior segments, ca 0.7 times as wide as the margin of metasternite, corresponding to a median peg on the posterior margin. Glandular pores clustered on the posterior part of the metasternites, in a short subtriangular area in the anterior part of the trunk, in two paired areas of fewer pores in the posterior part of the trunk. Procoxa 1.2–1.5 times as long as the metacoxa, both sclerites without glandular pores. Legs of the first pair slightly smaller than the subsequent ones. Leg claws simple, uniformly bent; a pair of accessory spines, the anterior one reaching approximately the midlength of the pretarsus, the posterior one distinctly shorter.
Ultimate leg-bearing segment (Figs 2,11). Pleuropretergite entire, lacking sutures or sulci or notches, ca 2.6 times as wide as long. Metatergite subtrapezoid, ca 1.1 times as wide as long, lateral margins convex and distinctly converging posteriorly, posterior margin convex. Presternite ca 8 times as wide as long, not medially constricted. Metasternite subtrapezoid, ca 1.7 times as wide as long, lateral margins slightly convex and converging backwards, posterior margin almost straight; setae uniformly sparse. Coxopleuron ca 2.0 times as long as wide, ca 1.5 times as long of the metasternite, anteriorly reaching the posterior margin of the presternite; short, dense setae on the postero-mesal part of the ventral side. Coxal organs of each coxopleuron opening through 8 pores approximately aligned into an elongate pouch covered by the metasternite and running along the lateral margin of the latter. The telopodite ca 13 times as long as wide, ca 1.5 times as long and 1.2 times as wide as the penultimate telopodite; 6 articles, gradually narrowing towards the tip; all articles covered dorsally with sparse longer setae, ventrally with dense shorter setae. Pretarsus in the shape of a pointed claw, similar to that of the preceding legs but only half the length of the penultimate, with two accessory spines, the posterior much longer than the anterior one.
Postpedal segments ( Fig. 11 View FIGURES 6 – 12 ). Genital pleurosternite entire, ca 2 times as wide as long, ca 1.5 times as long as the exposed part of the intermediate sternite. Gonopods in the shape of a short slightly bilobed lamina. Anal organs and pores indistinct.
Difference in an adult male. One of the syntypes, from Sevilla, Granada, 36 mm long, with 57 leg pairs ( Fig. 12 View FIGURES 6 – 12 ).
Ultimate leg-bearing segment ( Fig. 12 View FIGURES 6 – 12 ). Telopodite ca 7.5 times as long as wide, ca 1.3 times as long and 2.0 times as wide as the penultimate.
Postpedal segments ( Fig. 12 View FIGURES 6 – 12 ). Genital sternite ca 2 times as long as the exposed part of the intermediate sternite; complete sutures between pleurites and sternite, converging anteriorly to a point covered by the intermediate sternite. Gonopods biarticulated, well separated, with a subconic penis in between.
Variation. The body length was up to 51 mm in females (n=19) and up to 46 mm in males (n=10).
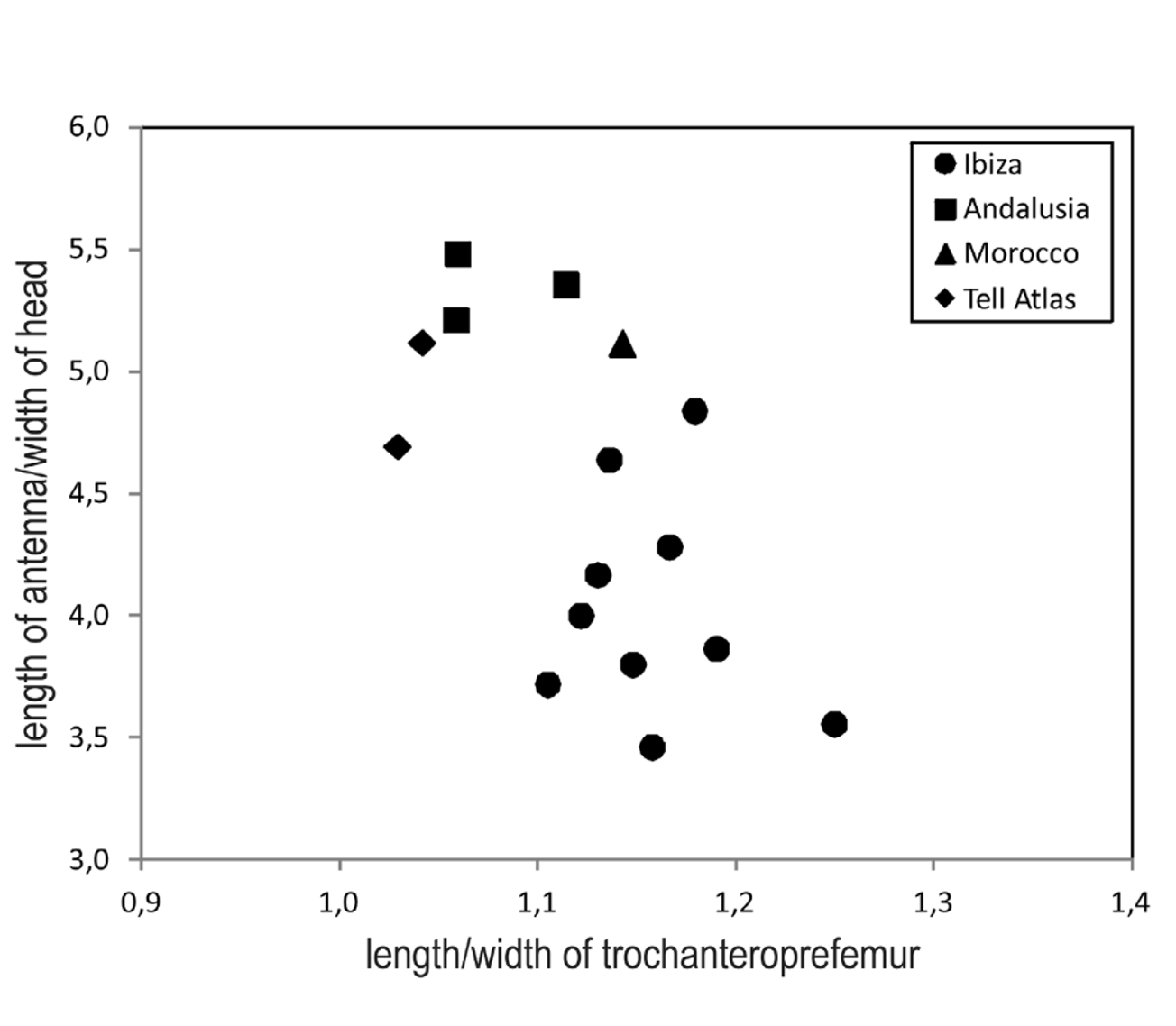
The number of legs varies in the range of 57–71 pairs in females (n=20) and 57–67 pairs in males (n=9), with a still lower value of 55 reported in the literature ( Brolemann 1932). Besides intra-population variability, some geographical variation has been detected: in particular, the numbers found in Ibiza (65–71 in females, n=5; 65–67 in males, n=4) are higher than those found in Andalusia (57–65 in females, n=11; 57–61 in males, n=6), the difference in females being statistically significant (Kolmogorov-Smirnov test: p<0.01); the specimens from African localities are too few to allow a comparison, but the numbers found (59–61 in 3 females) are within the range recorded in Andalusia.
Some geographical variation has been found also in the relative elongation of the antennae and the forcipular segments ( Fig. 13 View FIGURE 13 ). In particular, the specimens from Andalusia have on average relatively longer antennae and slightly less elongated forcipules than those from Ibiza, as suggested by the statistically significant differences in the elongation of the antennae (antennal length/head width: mean±standard deviation = 5.35± 0.14 in Andalusia, 4.00± 0.44 in Ibiza; Mann-Whitney test U=0.00, p=0.14, n=3, 8), the forcipular coxosternite (length/width of the ventrally exposed surface: mean±standard deviation = 0,82± 0.01 in Andalusia vs. 0.91± 0.04 in Ibiza; U=0.00, p=0.006, n=4, 8) and the forcipular trochanteroprefemur (length/width in ventral view: mean±standard deviation = 1.08± 0.03 in Andalusia vs. 1.17± 0.04 in Ibiza, U=0.00, p=0.014, n=3, 8). Instead, no variation has been found in the relative elongation of the head (length/width: U=18.0, p=0.770, n=8,5).
Taxonomic history. Geophilus hispanicus was described by Meinert (1870) upon a series of specimens of both sexes from Southern Spain. It was cited under the same generic combination by subsequent authors, sometimes including it in the subgenus Geophilus ( Verhoeff 1896; Attems 1903). Based on specimens from North-western Africa, Brolemann (1925, 1932, 1947) recognised that G. hispanicus deserved a distinct genus and therefore introduced Algerophilus for it. This action was overlooked by Attems (1929) and most of the later authors. Instead, G. hispanicus was assigned to either Tuoba or its synonym Nesogeophilus ( Demange 1963; Sammler et al. 2006). However, the true identity of the species remained poorly understood (e.g., Jeekel 2003).
Notes on synonymy. Nesogeophilus mateui was described by Machado (1953) on a single male from Southern Spain, but the species was completely neglected by later authors and its identity remained unassessed. Based on the original description and illustrations, N. mateui agrees with G. hispanicus , even in the traits which are recognised here as diagnostic of Algerophilus with respect to all other genera, including the shape and features of the forcipular coxosternite and the forcipules, the arrangement of the coxal pores, and the absence of anal pores. The only apparent difference between N. mateui and G. hispanicus is in the elongation of the head, which is described as about as long as wide in N. mateui whereas it is actually slightly longer than wide in A. hispanicus ( Meinert 1870; Fig. 1 View FIGURES 1 – 5 , our obs.). However, after examining specimens collected from near the type locality of N. mateui (see Specimens examined, and Fig. 13 View FIGURE 13 ), and therefore confidently recognisable as representative of this nominal species, we did not find any difference in the elongation of the head with respect to specimens from other localities.
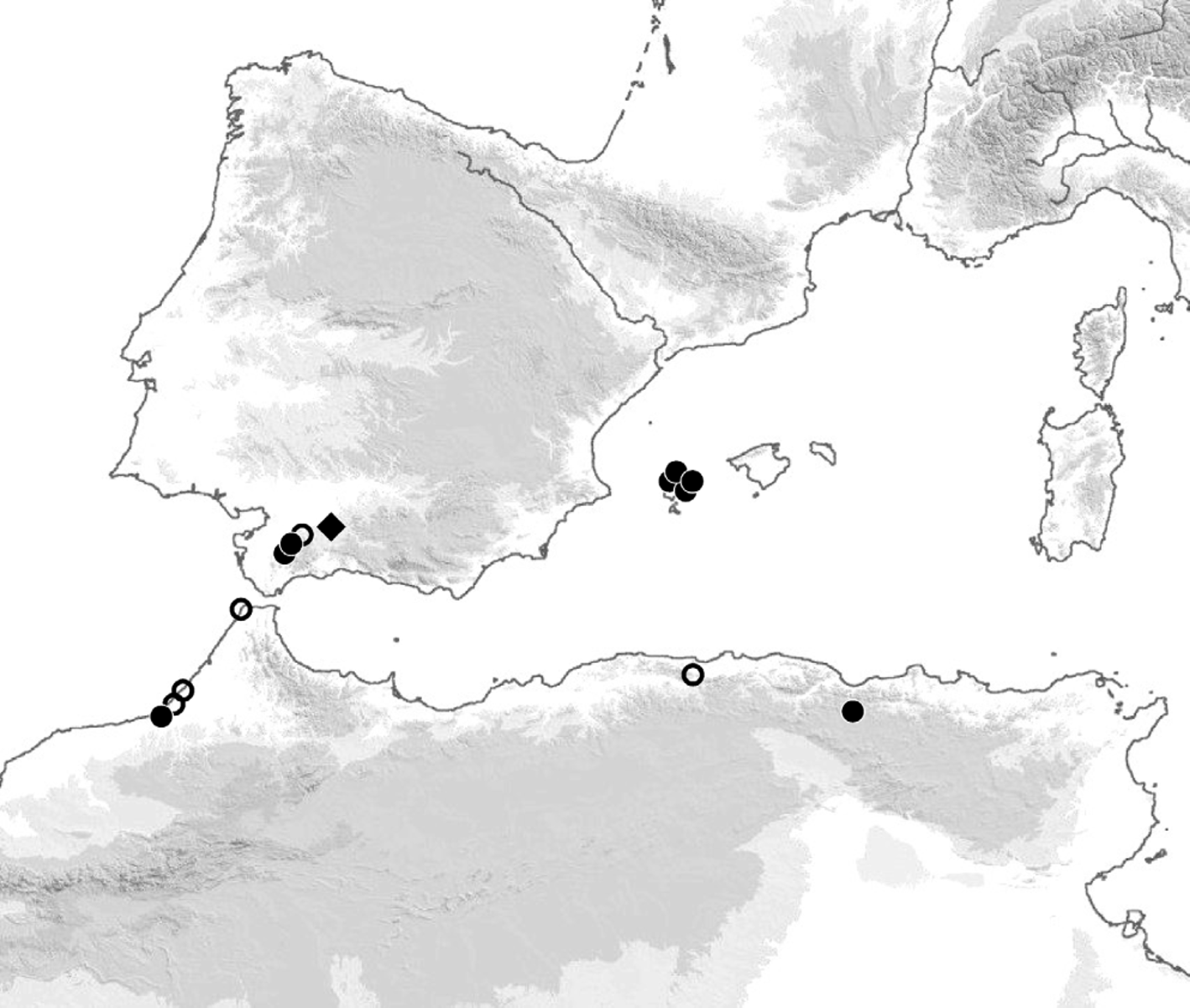
Geographical distribution. Taking into account all published and new records ( Fig. 14 View FIGURE 14 ), A. hispanicus has been recorded from a total of 15 localities in the most Western part of the Mediteranean region, i.e. in Andalusia ( Spain), the island of Ibiza ( Spain) and the northwestern part of Africa ( Morocco and Algeria).
The provenance of the syntypes of G. hispanicus is imprecisely known (it is given as “Sevilla, Granada ” both on the labels and in the published original description), nevertheless all records from the Iberian peninsula are most probably from the Baetic Mountains. Besides “ Granada, Sevilla” ( Meinert 1870), they include: Benaojan ( Machado 1953); Ubrique-Benaocaz, West of Benaocaz (new records). The records from Ibiza are from four coastal localities: Cala Olivera, Cap des Rubio, Can Codolar, La Joya ( Sammler et al. 2006). All four records from Morocco are from along the Atlantic coastal plain: Oued Fouarat near Kenitra, Oued Ykem, Tanger ( Brolemann 1932, 1947); Oued Cherrat (new record). The two records from Algeria are from in or near the Tell Atlas: Tipasa ( Brolemann 1925); Sétif (new record).
Habitat. Some information about environmental conditions is known only for a few sites. All Andalusian and Algerian records are from inland localities, up to about 50 kilometres from the sea coast, at least some of them in the mountains. Conversely, the Moroccan and Ibizan records are from coastal or sub-coastal localities, but they are referred to riverbeds, pine woods, juniper and heath scrubs, none to truly littoral habitat ( Brolemann 1932, Sammler et al. 2006).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Algerophilus hispanicus ( Meinert, 1870 )
| Bonato, Lucio, Voigtländer, Karin & Minelli, Alessandro 2012 |
Tuoba hispanica:
| Sammler et al. 2006 |
Nesogeophilus hispanicus:
| Demange 1963 |
Algerophilus hispanicus:
| Brolemann 1925 |
Geophilus (Geophilus) hispanicus:
| Verhoeff 1896 |
Geophilus hispanicus:
| Meinert 1870 |