
Pothea carpinteroi, Gil-Santana, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4778.3.2 |
|
publication LSID |
lsid:zoobank.org:pub:19CF6FB8-6018-4335-B462-62E8AD843C14 |
|
DOI |
https://doi.org/10.5281/zenodo.3856770 |
|
persistent identifier |
https://treatment.plazi.org/id/03CF87B7-C10E-ED38-FF7C-799C3CEDBF53 |
|
treatment provided by |
Plazi |
|
scientific name |
Pothea carpinteroi |
| status |
sp. nov. |
Pothea carpinteroi sp. nov.
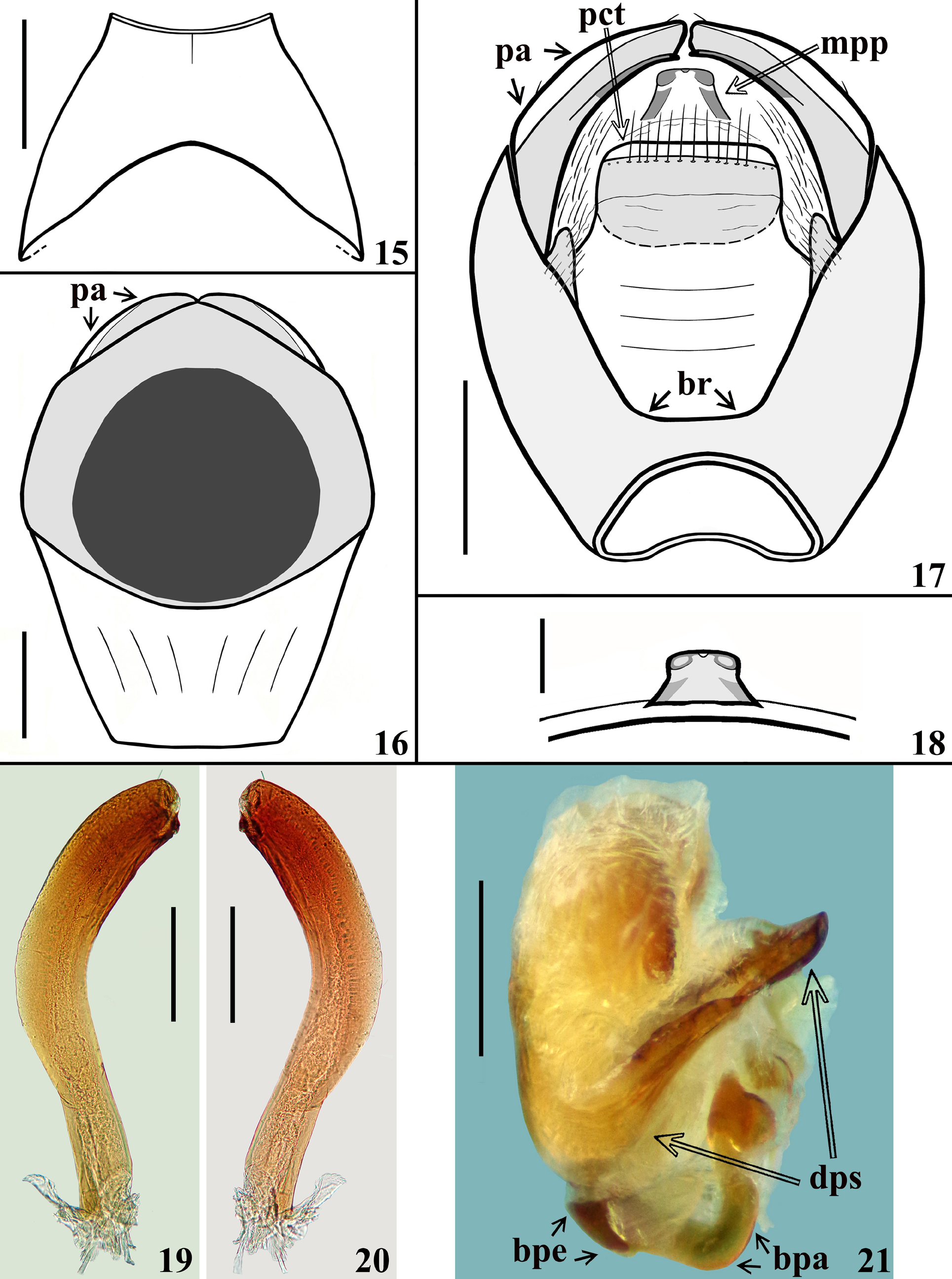
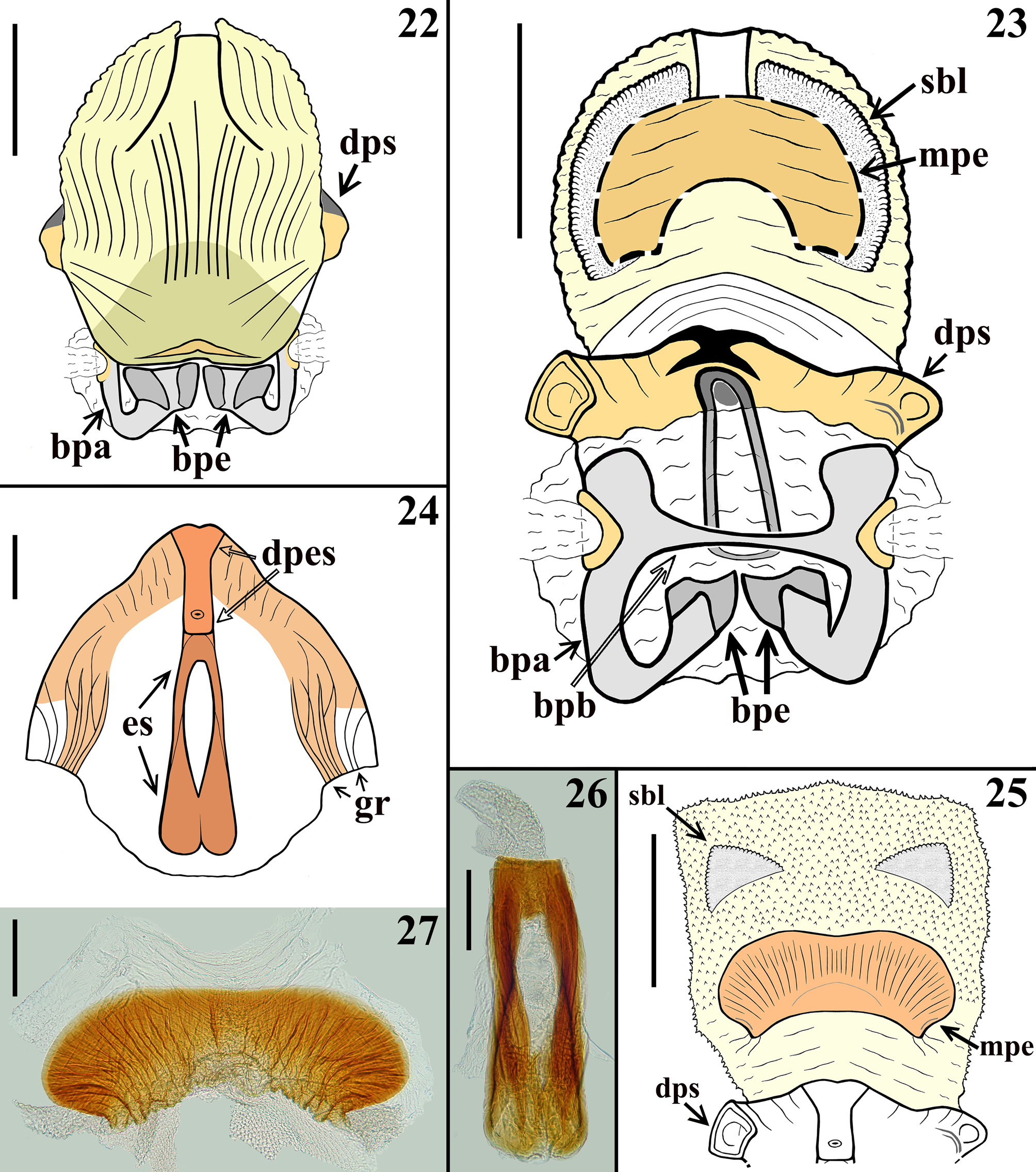
( Figs. 7–27 View FIGURES 7–10 View FIGURES 11–14 View FIGURES 15–21 View FIGURES 22–27 )
Diagnosis. The male most closely resembles that of Pothea jaguaris . The two species can be readily separated by the following characteristics: 1 – in P. carpinteroi sp. nov. the ratio between the total length (including collum) of the head is approximately twice the value of the maximum width across eyes of the head, while among males of P. jaguaris , it is around 1.6; 2 – height of the middle third of anteocular portion (maximum distance between its upper and lower margins) larger in P. carpinteroi sp. nov.; 3 – ocellar tubercle more prominent in P. carpinteroi sp. nov.; 4 – a small rounded tubercle on the ventral portion of the head (gula) anteriorly almost always present in P. jaguaris and absent in P. carpinteroi sp. nov.; 5 – longitudinal sulcus absent on anterior two-thirds of fore lobe of pronotum in P. carpinteroi sp. nov., clearly present in P. jaguaris , in which it can be somewhat shallow or deep enough to form a pair of protuberant paramedial lobes; 6 – prongs of scutellum widely separated at the base and convergent towards their apices in P. carpinteroi sp. nov. and more proximate at the base and running parallel to each other in P. jaguaris ; 7 – fore and middle trochanters and femora with papillae in P. carpinteroi sp. nov., which are absent in P. jaguaris ; 8 – pygophore: the bridge between anterior and posterior genital openings narrower in P. jaguaris ; 9 – midlateral portions of dorsal phallothecal sclerite with more grooves in P. jaguaris ; 10 – arms of endosomal struts more extensively united at basal portion and with their mid portions enlarged laterally in P. jaguaris ; 11 – median process of endosoma large, arcuate, diffusely finely striated in P. carpinteroi sp. nov. and small, subquadrate, with a central sclerotized structure in the shape of an “X” or a “T” in P. jaguaris .
Description. Male. COLORATION: General coloration yellowish-orange to somewhat yellow-brownish, with blackish to brownish markings or portions ( Figs. 7–14 View FIGURES 7–10 View FIGURES 11–14 ). Head ( Figs. 9–12 View FIGURES 7–10 View FIGURES 11–14 ) mostly blackish ventrally and yellowish to yellowish orange dorsally, especially between antennifers, eyes, around ocellar tubercle, on upper portion of maxillary plate, clypeus or only its proximal portion (paratype and holotype, respectively), almost entire labrum in paratype, anteromedian portion (holotype) or entire dorsal surface (paratype) of collum; in the paratype, yellowish brown medioventrally, from approximately the level of antennifers to collum, extending somewhat more laterally between eyes; ocellar tubercle mostly blackish, ocelli yellowish and surrounded anteriorly and posteriorly by nar- row reddish stripes; paratype with a subtle reddish tinge on median portion of clypeus, where it is denticulate. Labium: segment II (first visible) brownish to brownish black; segment III–IV yellowish to yellowish brown ( Fig. 12 View FIGURES 11–14 ). Antennal segments I–II generally blackish brown; base of the segment I pale; others absent. Thorax: fore lobe of pronotum mostly blackish, except the central portion of anterolateral angles which are slightly paler and the area just above the medial fovea of transverse furrow, which is yellow orange (holotype) ( Figs. 7, 9 View FIGURES 7–10 ) or with the anterolateral angles, lateral portions, except the posterior half of lateral margin, a rounded pale area around and including the medial impression of fore lobe, median portion of transverse sulcus, medial fovea and its margins, yellowish orange (paratype) ( Figs. 8, 10 View FIGURES 7–10 ); hind lobe of pronotum yellowish orange with a pair of suboval spots on central portion, which are quite larger in holotype, continuous with the blackish coloration of fore lobe, medially, reaching the posterior border and completely (holotype) ( Figs. 7, 9 View FIGURES 7–10 ) or shortly (paratype) united in the region of the median sulcus, below the medial large fovea of transverse furrow ( Figs. 8, 10 View FIGURES 7–10 ); lateral border (less extensively in paratype) blackish. Scutellum blackish ( Figs. 7–10 View FIGURES 7–10 ). Pleurae and sterna mostly blackish; posterior half of propleura yellowish brown; stridulatory sulcus generally paler. Legs: coxa and trochanters yellowish to yellowish brown; fore femora mostly blackish, narrowly (holotype) or more extensively (paratype) yellowish at basal portion and with a moderately wide subapical yellowish ring ( Figs. 7–10 View FIGURES 7–10 ); middle femora yellowish, with a larger (holotype) or narrower (paratype) submedian ring and apex blackish ( Figs. 7–10 View FIGURES 7–10 ); hind femora mostly pale yellowish with a moderately large (holotype) ( Fig. 13 View FIGURES 11–14 ) or narrower submedian distal dark ring (paratype) ( Fig. 8 View FIGURES 7–10 ) and apices blackish ( Figs. 7–8, 10 View FIGURES 7–10 , 13 View FIGURES 11–14 ); tibiae mostly blackish to blackish brown ( Figs. 7–8 View FIGURES 7–10 , 13 View FIGURES 11–14 ); dorsal portion of base of middle and hind tibiae slightly paler; fore tibiae paler at approximately the submedian proximal half; middle and hind tibiae with a pair of subbasal pale spots on inner and outer surfaces (holotype); in the paratype the spots on middle tibiae are somewhat more extensive and in hind tibiae there is a large submedian proximal pale ring which is approximately equivalent to one third to one fourth of the length of the segment ( Figs 7–8 View FIGURES 7–10 ). Tarsi generally yellowish to yellowish brown; fore and middle third tarsal segments darkened, except at extreme base and apex, which are paler; hind third tarsal segment somewhat darkened at approximately distal two-thirds. Hemelytra almost completely blackish, except for a basolateral yellowish orange spot on corium ( Figs. 7–10 View FIGURES 7–10 ), which is larger and has a subtle reddish tinge in posterior half in the paratype ( Figs. 8, 10 View FIGURES 7–10 ); areas subjacent to clavus and basal portions of cells of membrane somewhat paler ( Figs. 7–10 View FIGURES 7–10 ). Connexivum pale to pale yellow and clearly alternating pale and dark areas ( Figs. 7–8 View FIGURES 7–10 , 13–14 View FIGURES 11–14 ); blackish marked on: distal margin of segment II; approximately distal third to distal half of lateral and medial portions of segments III–VI in the holotype and approximately the distal fourth in the paratype, respectively; median distal portion of segment VII darkened. Sternite II blackish; sternites III–VI yellow brownish with large basal transverse blackish bands, continuous with a large lateral blackish longitudinal band, which borders the connexivum margin ( Figs. 13–14 View FIGURES 11–14 ); the connexival blackish markings are continuous with this lateral band too ( Figs. 13–14 View FIGURES 11–14 ); small pale irregular spots in the lateral band, in the basal portions of segments IV–VI in the paratype; in the holotype, similar, but smaller, almost imperceptible pale spots only on basal portions of segments V–VI; in the paratype, the transverse bands are paler at midline on the sternites IV–V and interrupted on sternites VI–VII at midline ( Fig. 14 View FIGURES 11–14 ). Sternite VII almost completely blackish at median portion, with a pale spot at midline, just above pygophore (holotype) ( Fig. 13 View FIGURES 11–14 ); in paratype, the portion above pygophore mostly yellowish to yellowish orange, with a pair of basal transverse bands, which are narrower, shorter, not reaching the lateral band, somewhat curved, obliquely directed backwards, far from intersegmental suture laterally; the lateral band evident, but becoming narrower and ending approximately at the level of anterior margin of pygophore ( Fig. 14 View FIGURES 11–14 ). Exposed portion of pygophore generally paler at lateral and posterior borders and darkened to blackish on central and basal portion ( Figs. 13–14 View FIGURES 11–14 ). STRUCTURE: Integument mostly shiny and smooth. Head ( Figs. 9–12 View FIGURES 7–10 View FIGURES 11–14 ) elongated, almost as long as pronotum (excluding collum); anteocular portion approximately twice and a half longer than postocular portion (excluding collum); ratio between the total length (including collum) and maximum width across eyes of the head: 2.17–2.00 (holotype / paratype); minimum distance between eyes in dorsal view (synthlipsis) somewhat longer than the width of each eye; height (maximum distance between upper and lower margin of the head) larger on middle third of anteocular portion ( Fig. 12 View FIGURES 11–14 ); antennifers distant from anterior margin of eyes; eyes prominent, rounded in dorsal view ( Fig. 11 View FIGURES 11–14 ), suboval in lateral view ( Fig. 12 View FIGURES 11–14 ); transverse sulcus barely marked at lateral portions, not evident at median portion ( Fig. 11 View FIGURES 11–14 ); ocellar tubercle prominent, undivided, ocelli rounded, distance between them the same as the diameter of each ocelli ( Figs. 11–12 View FIGURES 11–14 ); antenna inserted somewhat proximal to midpoint between anterior margin of eyes and apex of the head ( Figs. 11–12 View FIGURES 11–14 ); scape surpassing apex of head by somewhat less than its distal third, somewhat curved and enlarged towards apex, shorter than pedicel and anteocular portion of head; pedicel straight; remaining antennal segments absent. Clypeus elongate, denticulate in lateral view ( Fig. 12 View FIGURES 11–14 ). Labium thick, length ratio between the labial (visible) segments 4.1–3.4:1.8–1.7:1.0; segment II (first visible) straight, approximately twice longer than the segment III, also longer than the others together, its apex approximately at level of the posterior margin of eyes; segment III somewhat enlarged at distal half; segment IV, shorter, tapering ( Fig. 12 View FIGURES 11–14 ). Thorax ( Figs. 7–10 View FIGURES 7–10 ): integument smooth, shiny; collar thin; anterolateral angles rounded and small; fore lobe rounded on anterior and lateral margins, shorter and narrower than hind lobe; midlongitudinal furrow on fore lobe represented by a shallow median small depression just above a fovea, at median portion of transverse furrow, the latter interrupting the midlongitudinal furrow, which on hind lobe is thin but with somewhat irregular margins and shortly exceeding the basal half of the hind lobe; transverse furrow distinct, carinulate, interrupted medially by the aforementioned median fovea, continuing laterally, on propleura, where it is formed by somewhat larger and deeper depressions, forming a somewhat curved lateral furrow which ends at short distance anteriorly to the posterior margin of propleura; posterolateral furrows of pronotum distinct and formed by a series of shallow punctuations; humeral angles rounded. Scutellum with a shallow median depression; prongs widely separated at the base and convergent towards their apices. Supracoxal lobes of propleura somewhat prominent, those of meso and metapleura not; integument of anterior portion of propleura with few deep irregular punctuations; integument of mesopleura mostly smooth; metapleura coarsely rugous, with several linear subparallel irregular ridges, superior margin thickened and curved. Prosternum wider on approximately anterior half in which there is a pair of short, rounded, lateral processes, directed forward and narrowing posteriorly; on posterior half, the prosternum forms a cylindrical median process, which surpasses fore coxae and reaches mesosternum, with its median portion mostly occupied by the stridulitrum. Mesosternum anteriorly to middle coxa with smooth integument, a pair of rounded small depressions medial to the superior margins of middle coxal cavities; between middle coxa, a subrectangular moderately elevated area with integument marked by few shallow transverse lines. Metasternum short. Fore coxae close, separated by a distance shorter than approximately half the width of each of them; middle and hind coxae distant from each other by a distance approximately equivalent to 1.5 and the same width of each of them, respectively. Fore and middle trochanters with a median row of papillae on distal half; fore trochanter with an additional similar row of papillae, anteriorly to the median row. Fore and middle femora somewhat thickened ( Figs. 7–10 View FIGURES 7–10 ), with their maximum width at approximately mid and submedian distal portions respectively; hind femora slender, somewhat thickened subapically ( Figs. 7–8 View FIGURES 7–10 , 13 View FIGURES 11–14 ). On fore and middle femora, a median ventral shallow and thin crest, which are larger basally, becoming narrower towards apices; on this crest there are papillae and a ventral fringe of pale setae; papillae more numerous at basal portion. Fore and middle tibiae straight, hind tibia slightly curved at distal half ( Figs. 7 View FIGURES 7–10 , 13 View FIGURES 11–14 ); fore tibiae conspicuously thicker at apex, in which the anterior margin is prominent; mid and hind tibiae progressively less thicker at apex; spongy fossa on fore and middle tibiae moderately developed. All tarsi slender, three-segmented. Hemelytra not reaching tip of abdomen, generally dull; on extreme base of dorsal surface, laterally, and on lateral portion, basally, moderately shiny ( Figs. 7–8 View FIGURES 7–10 ). Abdomen: connexivum with posterolateral angle between segments II and III somewhat prominent; dorsal connexival integument somewhat rugous, except on the lateral margin, in which the integument is smooth. Sternites with smooth, shiny integument; sternite II narrower than the following segments, median portion somewhat elevated and with the integument slightly rugous; sternites II and III separated by canaliculae; other intersternite furrows more evident in median portion, as a thin line, and almost imperceptible laterally; the furrow between segments VI and VII somewhat more marked and depressed laterally to midline. Segment VIII not visible externally, sclerotized on ventral portion, which becomes wider towards posterior margin; latter curved ( Fig. 15 View FIGURES 15–21 ); dorsal portion membranous and narrower; spiracles on dorsal margin of ventral portion. Male genitalia ( Figs.16–27 View FIGURES 15–21 View FIGURES 22–27 ). Genital capsule, in ventral and lateral views: exposed portion of pygophore subhexagonal ( Fig. 16 View FIGURES 15–21 ) and rounded, respectively, integument glabrous, smooth and shiny; not pigmented in the ventral non-exposed portion; in dorsal view ( Fig. 17 View FIGURES 15–21 ): between anterior and posterior genital openings, a narrow well-sclerotized dorsal (transverse) somewhat curved bridge (br); membranous areas of posterior genital opening smooth; proctiger (pct) subrectangular, posterior margin almost straight, slightly curved laterally, with a subapical row of long straight setae. Medial process of pygophore (mpp) esclerotized, apical half subquadrate in anterior view, apical margin almost straight, slightly curved ( Fig. 18 View FIGURES 15–21 ). Parameres (pa) mildly exposed when genital capsule is in situ or in ventral view ( Fig. 16 View FIGURES 15–21 ); their apices in contact in resting position ( Fig. 17 View FIGURES 15–21 ). Parameres symmetrical, elongated, curved at approximately middle third; apex truncated, with a short subapical tooth in inferior margin; mostly glabrous, with a few scattered short setae on upper surface ( Figs. 17, 19–20 View FIGURES 15–21 ). Phallus ( Figs. 21–23 View FIGURES 15–21 View FIGURES 22–27 ): articulatory apparatus with basal plate extension (bpe) much shorter than basal plate, the latter with moderately short and curved basal plate arms (bpa), connected by a narrow basal plate bridge (bpb). Dorsal phallothecal sclerite (dps) symmetrical, enlarged to the apex, sinuous in center of anterior margin and more pronouncedly sinuate laterally to anterior margin; midlateral portions with few grooves (gr) ( Figs. 21 View FIGURES 15–21 , 23–24 View FIGURES 22–27 ). Endosomal struts (es) formed by a pair of parallel arms, nearly straight at mid portion, larger at basal portion, united at base and apex ( Figs. 24, 26 View FIGURES 22–27 ), which is continuous with the dorsal phallothecal sclerite-endosomal struts fusion (dpes) ( Fig. 24 View FIGURES 22–27 ); the latter is shorter than the endosomal struts (es) and somewhat enlarged at approximately distal third, toward apex of apical margin of dorsal phallothecal sclerite (dps) ( Fig. 24 View FIGURES 22–27 ). Endosoma wall longitudinally striated on basal portion, ventrally ( Fig. 22 View FIGURES 22–27 ), smooth basally, and mostly densely minutely, spiny, with a pair of flat lateral small subapical lobes (sbl) ( Figs. 23, 25 View FIGURES 22–27 ). Median process of endosoma (mpe) large, arcuate, finely striated ( Figs. 23, 25, 27 View FIGURES 22–27 ). VESTITURE: Body mostly glabrous. Head: sparse curved short or somewhat longer, pale to yellowish setae on distal portion of clypeus, numerous on labrum; some longer similar setae scattered on anterior and lateral portions of base of first visible labial segment; last labial segment with few scattered erect, thin, pale setae. Antennal segments I–II (others absent) generally covered by very short, adpressed, pale, curved, thin setae and long, fine, erect, stout, yellowish to yellowish brown numerous setae; first segment almost bare in mid-dorsal region, with only a few scattered adpressed setae, the erect setae inserted on its lateral and ventral portions and as long as or somewhat longer than the transverse width of segment; second antennal segment with the erect setae two to almost three times as long as the width of the segment, sparse on dorsal surface and very numerous on the remaining surfaces; second segment also covered by numerous oblique, stout, short, straight to somewhat curved numerous yellowish brown to brownish setae on all surfaces of the segment. Thorax: pronotum, scutellum, pleurae and hemelytra glabrous; thoracic sterna mostly glabrous; on prosternum, laterally to stridulitrum, a pubescence formed by very short, thin, golden setae; borders of posterior prolongation of prosternum with numerous thin longer setae; some groupings of pale thin setae on anterior margin of mesosternum, laterally to the prolongation of prosternum; a patch of yellowish curved setae on medial wall of middle coxal cavity. Legs: coxa mostly glabrous, with a single long somewhat curved golden seta implanted on apico-medial portion; all trochanters with a single long, somewhat curved pale setae implanted on basal portion, ventrally; fore trochanter with two rows of numerous yellowish to golden, curved, thin setae on approximately distal two thirds of ventral surface, in the same portion where there are papillae; on middle and hind trochanters these setae form a group inserted on approximately two thirds to distal half of ventral surface, respectively, together with papillae in the middle trochanter and somewhat less numerous in hind trochanter. Femora mostly glabrous; fore femora with a single elongated somewhat curved, pale, ventral, subbasal setae; all femora with an apicoventral pair of similar setae; middle femora with additional sparse somewhat shorter, apical setae, on lateral and dorsal portions; fore and middle femora with a ventral fringe of numerous decumbent, curved, thin, pale setae inserted on the median ventral crest, in the same region where there are papillae; the ventral fringe is larger and the setae are more numerous in basal portion, becoming progressively narrower and less numerous, respectively, towards apical portion; hind femora with a few golden setae on basal portion of the segment, ventrally. Tibiae with a mid ventral fringe of short, straight, somewhat stouter, yellowish to golden or more darkened setae; on fore and middle tibiae the fringe are narrower; at approximately distal third, all tibiae generally covered by golden to darkened setae around the segment, which become more numerous and somewhat longer towards apex of segment. Tarsi covered with numerous yellowish and golden setae, which are longer on ventral surface. Abdomen glabrous. MEASUREMENTS: (holotype and paratype, in mm): total length: to tip of abdomen 16.1–14.5; to tip of hemelytra 15.8–14.3; head: length: including collum 3.9–3.2; excluding collum 3.4–2.9; maximum height (maximum distance between upper and lower margin of the head) 1.4–1.2; length of anteocular portion (measured on lateral view) 2.0–1.7; length of postocular portion (measured on lateral view), including collum 1.2; excluding collum 0.8–0.6; width across eyes 1.8–1.6; minimum interocular distance between eyes, dorsal view (synthlipsis) 0.6; width of eye, dorsal view 0.5; length of eye, dorsal view 0.6–0.5; maximum height of eye, lateral view 0.9–0.8; distance between external margins of ocellar tubercle 0.6–0.5; distance between ocelli 0.15; maximum width of ocellus: 0.15; length of ocellar tubercle on midline 0.5; antennal segments: I (scape) 1.2–1.3; II (pedicel) 2.5; others absent; labial segments: II [first visible] 2.5–2.0; III 1.1–1.0; IV 0.6. Thorax: pronotum: fore lobe: length 1.4–1.3; maximum width 2.8–2.5; hind lobe: length 1.7–1.5; maximum width 4.1–3.7; scutellum: length 1.5–1.4; width at base 1.8–1.7; minimum distance between prongs 0.3–0.25. Fore legs: length of femur 3.6–3.0, maximum width (at midportion) 0.7–0.6; length of tibia 3.7–3.2; length of spongy fossa 0.6; length of tarsus 1.2–absent; middle legs: length of femur 3.3–3.2, maximum width (at submedian distal portion) 0.6; length of tibia 3.5–3.1; length of spongy fossa 0.7–0.6; length of tarsus absent–1.1; hind legs: length of femur 4.7–4.1, width at midportion 0.4, maximum width (at subapical portion) 0.5; length of tibia 5.4–4.8; length of tarsus 1.5–1.4. Abdomen: length 7.5–7.1; maximum width 5.0–4.4.
Distribution. Mexico.
Etymology. The new species is named in honor of the late Dr Diego José Carpintero, from Argentina, for his great contribution to the study of Reduviidae , particularly to the Ectrichodiinae , including the genus Pothea .
Type material. MEXICO, Quintana Roo, Cobá , v. 1979, Fernandéz, J. leg., male holotype, 1 male paratype ( MNRJ) .
Material of other related specimens examined:
Parapothea jaguaris Carpintero, 1980 . Female holotype: ♀ [handwritten] / HOLOTYPUS [printed] / P. j. 1 [handwritten] [red label] // Feb. - 952 [vertical line at left side] / BOLIVIA / Do Cochabamba / O?ia Chapare / Villa ??al. Román / uso????. aet. Coll. Martínez [handwritten label], [opposite side of same label]: Pothea ♀ / sp. nov. / P. wygodzinsky [handwritten] / A. MARTINEZ- DET. 1953 [printed, with exception of “53”, which was handwritten] //
Parapothea / jaguaris / N. S. / [printed horizontal line] / Det. Dr. Carpintero [printed] ( MACN). Additional specimens: FRENCH GUIANA, Nourages [04º 52’ N– 52º 41’ W], ix.2009, window trap, leg. SEAG (3 males) GoogleMaps .
Pothea bivittata Champion, 1899 . Female holotype: [ Guatemala]: B.C.A.Rhyn. II. / Pothea / bivittata Ch [handwritten] / ♀ [handwritten] // Sp. figured. // Balheu, / Vera Paz. / Champion. // Type [rounded label with orange circle] // ♀ // Holo- / type [rounded label with red circle] // [QR code] / NHMUK 013588139 About NHMUK ( BMNH) .
Pothea halffteri Carpintero, 1980 . Male holotype: ♂ [handwritten] / HOLOTYPUS [printed] [red label] // AGO. [august].975 [vertical line at left side] / MEXICO / VERACRUZ / Los Tuxtlas / Bert – legit / Coll. Martínez [handwritten label] // Pothea / ( Pothea ) / halffteri N. S [handwritten] / [printed horizontal line] / Det. Dr. Carpintero [printed] ( MACN).
Pothea maculata Champion, 1899 . Female holotype: B.C.A.Rhyn. II. / Pothea / maculata [handwritten] / ♂ Ch [handwritten] // Sp. figured. // Jalapa, / Mexico. / Hoege. // Type [rounded label with orange circle] // ♀ // Holo- / type [rounded label with red circle] // [QR code] / NHMUK 013585369 About NHMUK ( BMNH) .
Pothea reyesi Carpintero, 1980 . Male holotype: ♂ [handwritten] / HOLOTYPUS [printed] [red label] // Pothea / ( Pothea ) / reyesi N. S. [handwritten] / [printed horizontal line] / Det. Dr. Carpintero [printed] // Jul.969 [vertical line at left side] / MEXICO / COLIMA / TECOMÁN / E. Martin – leg. / Coll. Martínez [handwritten label] ( MACN).
Comments. Inclusion of the new species in Pothea is in accordance with the characteristics assigned to species of this genus by Carpintero (1978), Dougherty (1995), Maldonado & Carpintero (1996), Gil-Santana (2014) and Gil-Santana et al. (2015).
The general structure and especially the coloration of P. carpinteroi sp. nov. seem more similar to P. jaguaris than other congeners ( Gil-Santana 2014) ( Figs. 28–36 View FIGURES 28–30 View FIGURES 31–36 ). Although differences in coloration were recorded between the two specimens of P. carpinteroi sp. nov. examined here, a larger variation was recorded in P. jaguaris , apparently because more specimens of the latter species were available ( Gil-Santana 2014). Even considering a potentially broader variability in P. carpinteroi sp. nov., the colour of this species seemed more similar to darker and more extensively marked specimens of P. jaguaris . Nevertheless, colour differences were not considered for diagnosing the species due to uncertain range of variation of both species, only the structural differences between P. carpinteroi sp. nov. and P. jaguaris , already recorded in the diagnosis provided above.
As discussed by Gil-Santana (2014), in Pothea , the longitudinal sulcus on the anterior lobe of the pronotum has been considered obsolete, reduced to a medial or discal fovea either well developed or weakly developed anteriorly in this genus ( Carpintero 1978, Carpintero & Maldonado 1996, Dougherty 1995), but always regarded as a valuable taxonomic characteristic to differentiate species in this genus as well as Ectrichodiinae genera in general ( Carpintero & Maldonado 1996, Dougherty 1995, Gil-Santana et al. 2015, Gil-Santana 2019). Although Gil-Santana (2014) had recorded some variation in the deepness of the sulcus on fore lobe in P. jaguaris , its presence or absence is usually constant for all species of Ectrichodiinae in general (e.g. Wygodzinsky 1951, Carpintero 1978, Carpintero & Maldonado 1996, Dougherty 1995, Gil-Santana 2019). Therefore, its absence in P. carpinteroi sp. nov. ( Figs. 9–11 View FIGURES 7–10 View FIGURES 11–14 ) when compared with its constant presence in P. jaguaris ( Fig. 29 View FIGURES 28–30 ) seems to be a good structural characteristic to separate them. It is noteworthy to mention the presence of papillae on the fore and middle trochanters and femora in P. carpinteroi sp. nov., and their absence in P. jaguaris . In the male genitalia the most conspicuous difference was observed between the median processes of endosoma (mpe): large, arcuate, and finely striated in P. carpinteroi sp. nov. ( Figs. 23, 25, 27 View FIGURES 22–27 ) and small, subquadrate, with a central sclerotized structure in the shape of an “X” or a “T” in P. jaguaris ( Figs. 33, 36 View FIGURES 31–36 ). Yet, additional differences in the male genitalia were noted: the bridge (br) between the anterior and posterior genital openings of the pygophore is narrower in P. jaguaris ( Figs. 17 View FIGURES 15–21 ; 31 View FIGURES 31–36 ); the midlateral portions of the dorsal phallothecal sclerite (dps) have more grooves (gr) in P. jaguaris ( Fig. 33 View FIGURES 31–36 ) than those of P. carpinteroi sp. nov. ( Fig. 24 View FIGURES 22–27 ); and the basal and mid portions of the arms of the endosomal struts (es) are respectively united for a shorter distance and nearly straight in P. carpinteroi sp. nov. ( Figs. 24, 26 View FIGURES 22–27 ), while they are united for a comparatively longer distance basally and enlarged laterally at mid portion in P. jaguaris ( Figs. 33–35 View FIGURES 31–36 ).
Although the medial process of the pygophore (mpp) is a structure frequently useful for taxonomic purposes, it was not taken into account because of the fact that in P. jaguaris , variation in the shape of this structure was recorded between males of the same population ( Figs. 32 View FIGURES 31–36 , A–B). In this species, the shape of the apex of the medial process of the pygophore (in anterior view) showed to be rounded to slightly subtriangular ( Gil-Santana 2014, Figs. 32 View FIGURES 31–36 , A), or almost straight ( Fig. 32 View FIGURES 31–36 , B). In the latter case, it would not be significantly different from that of P. carpinteroi sp. nov. ( Fig. 18 View FIGURES 15–21 ).
Male genitalic characters apparently have little utility in the taxonomy of Neotropical Ectrichodiinae ( Dougherty 1995, Carpintero & Maldonado 1996). Dougherty (1995) posited the explanation that there were so few sclerotized structures in the phallus of Ectrichodiinae that there were no apparent differences among the various genera, with just a pair of sclerotized plates at the distal tip of the inflated endosoma. Nevertheless, differences in the male genital structures have been documented in some Brontostoma species ( Ectrichodiinae ) (Gil-Santana et al. 2005, Gil-Santana & Baena 2009, Gil-Santana et al. 2013), three species of Pothea ( Gil-Santana 2014) , Pseudopothea paulai Gil-Santana, 2015 (Gil-Santana 2015) and Sinchocoris giupponii Gil-Santana, 2019 (Gil-Santana 2019), thus highlighting the value of this character system for taxonomic and systematic studies in the subfamily Ectrichodiinae .
The male genitalia of three species of Pothea studied by Gil-Santana (2014) revealed several differences, mainly in the shape of the parameres, the medial process of the pygophore, the dorsal phallothecal sclerite and endosomal struts, and the median process of the endosoma, which appeared to be useful in the taxonomy of this group. The dorsal phallothecal sclerite and endosomal struts were particularly noteworthy, since their shape and “design” showed to be invariable within species and seem very particular to each species ( Gil-Santana 2014).
Between P. carpinteroi sp. nov. and P. jaguaris , although the parameres and the medial process of the pygophore were not useful to differentiate these species, almost all other differences were recorded in the aforementioned structures (2014) i.e., the dorsal phallothecal sclerite (24; 33), endosomal struts ( Figs. 24, 26 View FIGURES 22–27 ; 33–35 View FIGURES 31–36 ), and the median process of endosoma ( Figs. 23, 25, 27 View FIGURES 22–27 ; 33, 36 View FIGURES 31–36 ).
Nevertheless, in spite of the general similarity in the coloration of P. jaguaris and P. carpinteroi sp. nov., the validity of the latter, taking into account the structural differences recorded between them, can be considered as being in accordance to previous concepts and observations about the taxonomic value of certain morphological characteristics among species of Pothea and even Ectrichodiinae in general.
Besides P. jaguaris and P. carpinteroi sp. nov., three other species of Pothea , P. halffteri ( Figs. 37–38 View FIGURES 37–44 ), P. maculata ( Figs. 39–42 View FIGURES 37–44 ), and P. reyesi ( Figs. 43–44 View FIGURES 37–44 ) also present a pair of suboval or rounded dark spots on the hind lobe of pronotum, a conspicuous similarity among them. Therefore, it is noteworthy to mention the main differences or characteristics of those species because all of them were described from Mexico too. They can be easily separated by the following set of characters. Pothea halffteri has a general coloration quite diverse, particularly the uniform pale connexivum and the entirely blackish legs ( Fig. 37 View FIGURES 37–44 ); the clypeus is not denticulate; the fore and middle femora are thinner, without ventral crests ( Carpintero 1980). Pothea maculata is smaller (total length 12 mm); the head is shorter than pronotum ( Fig. 39 View FIGURES 37–44 ); the general coloration is brighter ( Figs. 39–41 View FIGURES 37–44 ); hemelytra pale brownish with the coloration of the lateral portion of the coria paler, almost whitish and more extensive, reaching beyond the middle ( Fig. 39 View FIGURES 37–44 ); the connexivum pale, spotted only on inner side of segments III–VI ( Champion 1899) and faintly so on segment VII ( Fig. 39 View FIGURES 37–44 ); the basal dark markings on sternites are smaller, not reaching the median portion of them, and therefore, not forming bands ( Fig. 40 View FIGURES 37–44 ). Pothea reyesi ( Fig. 43 View FIGURES 37–44 ) has a general brownish pale coloration, with fewer dark markings or portions; the pair of black spots on the hind lobe of the pronotum are rounded and small; the hemelytra are mostly brownish and the connexivum is uniformly pale colored ( Carpintero 1980). When describing P. halffteri and P. reyesi, Carpintero (1980) considered them close to P. bivittata ( Figs. 45–46, 48–50 View FIGURES 45–51 ), a species recorded from Guatemala and Mexico ( Champion 1899, Dougherty 1995). Thus, it is necessary to point the main differences between P. carpinteroi sp. nov. and P. bivittata too. Firstly, the pattern of coloration of the pronotum is diverse from P. carpinteroi sp. nov. ( Figs. 7–10 View FIGURES 7–10 ): in P. bivittata most of the head and pronotum is rufo-testaceous or reddish ( Figs. 45–46, 48–50 View FIGURES 45–51 ); the pronotum has a broad median vitta, which extends onto the posterior lobe “(leaving two sinuous vittae on the anterior lobe and a large black patch on the disc of the posterior lobe)” ( Champion 1899; see his fig. 13, Tab. XIII) ( Fig. 45 View FIGURES 45–51 ) or an anterior pair of small dark spots on fore lobe, just below the anterolateral angles, and a pair of large dark suboval spots on central portion of the hind lobe ( Fig. 48 View FIGURES 45–51 ); secondly, the connexivum has complete alternating pale and dark areas in P. carpinteroi sp. nov. ( Figs. 7–8 View FIGURES 7–10 , 13–14 View FIGURES 11–14 ), while it is entirely rufo-testaceous or pale at its external margin in P. bivittata ( Champion 1899; Figs. 45, 48 View FIGURES 45–51 ); thirdly, other portions which are darkened to yellow-brownish in P. carpinteroi sp. nov. are completely or more extensively rufo-testaceous or reddish, respectively, in P. bivittata , such as the ocellar tubercle, scutellar elevated lateral portion and its processes, some spots on pleura; fourthly, the pale coloration of the coria is more extensive in P. bivittata , reaching beyond the middle ( Figs. 45, 48 View FIGURES 45–51 ). The midlongitudinal furrow on the fore lobe is represented by a shallow median small depression in P. carpinteroi sp. nov., which is deeper and larger in P. bivittata ( Fig. 48 View FIGURES 45–51 ). On one hand, some similarities between these two species are noteworthy such as the shape of the head in lateral view ( Figs. 12 View FIGURES 11–14 ; 50 View FIGURES 45–51 ); the clypeus denticulate in the male ( Figs. 12 View FIGURES 11–14 ; 49 View FIGURES 45–51 ); a similar pattern of darkened portions in the legs; fore and middle femora somewhat thickened ( Figs. 7–10 View FIGURES 7–10 ; 45–46, 48–50 View FIGURES 45–51 ); and the coloration of the sternites, with large basal transverse blackish bands, continuous with a large lateral blackish longitudinal band ( Figs. 13–14 View FIGURES 11–14 ; 46, 50 View FIGURES 45–51 ). The presence of dark suboval spots on central portion of the hind lobe in the male ( Fig. 48 View FIGURES 45–51 ) is a similarity shared not only with P. carpinteroi sp. nov. ( Figs. 7–10 View FIGURES 7–10 ), but also with P. halffteri ( Fig. 37 View FIGURES 37–44 ), P. maculata ( Fig. 39 View FIGURES 37–44 ), and P. reyesi ( Fig. 43 View FIGURES 37–44 ), as mentioned above. On the other hand, taking into account that P. bivittata is known from a male and a female only, although they seem to be different species based on the available scarce material, in case that more specimens are found in the future, the distinctness or eventually the conspecificity of them will be confirmed or revealed, respectively.
Pothea venatrix ( Figs. 52–55 View FIGURES 52–55 ) presents two spots on the fore lobe and two vittae on the hind lobe of the pronotum ( Hussey 1953) ( Fig. 52 View FIGURES 52–55 ) and was considered very close to P. bivittata by this author. Therefore, it is also necessary to point the main differences of P. venatrix in relation to P. carpinteroi sp. nov.: P. venatrix has a diverse general coloration, with the head mostly pale testaceous, excepting the apices of antenniferous tubercles which are “embrowned” ( Hussey 1953) ( Figs. 52–54 View FIGURES 52–55 ); clypeus not denticulate ( Fig. 54 View FIGURES 52–55 ); pronotum with less extensive dark markings as described above; the lateral portions of hind lobe of pronotum somewhat reddish ( Fig. 52 View FIGURES 52–55 ); femora and tibiae dark brownish, the former with basal portions and pale subapical spots ( Figs. 52–53 View FIGURES 52–55 ); connexivum largely yellowish on external margin, without alternating pale and dark areas ( Figs. 52–53 View FIGURES 52–55 ); reddish testaceous sternites, lateral longitudinal band running only from sternite III to the last, with irregular borders, the basal transverse dark markings on sternites are thin, forming a band only on segment VII, while on segments IV–VI they form incomplete bands which do not reach the lateral dark band and are variably interrupted at midline ( Fig. 53 View FIGURES 52–55 ).







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Ectrichodiinae |
|
Genus |