
Crassimarginatella kumatae ( Okada, 1923 )
|
publication ID |
https://doi.org/10.11646/zootaxa.4226.4.2 |
|
publication LSID |
lsid:zoobank.org:pub:758449A2-1B82-45E2-B6C4-8D83354C6CA7 |
|
DOI |
https://doi.org/10.5281/zenodo.5697210 |
|
persistent identifier |
https://treatment.plazi.org/id/03CF87E8-FFE0-FF8E-FF15-FC98FD982046 |
|
treatment provided by |
Plazi |
|
scientific name |
Crassimarginatella kumatae ( Okada, 1923 ) |
| status |
|
Crassimarginatella kumatae ( Okada, 1923) View in CoL
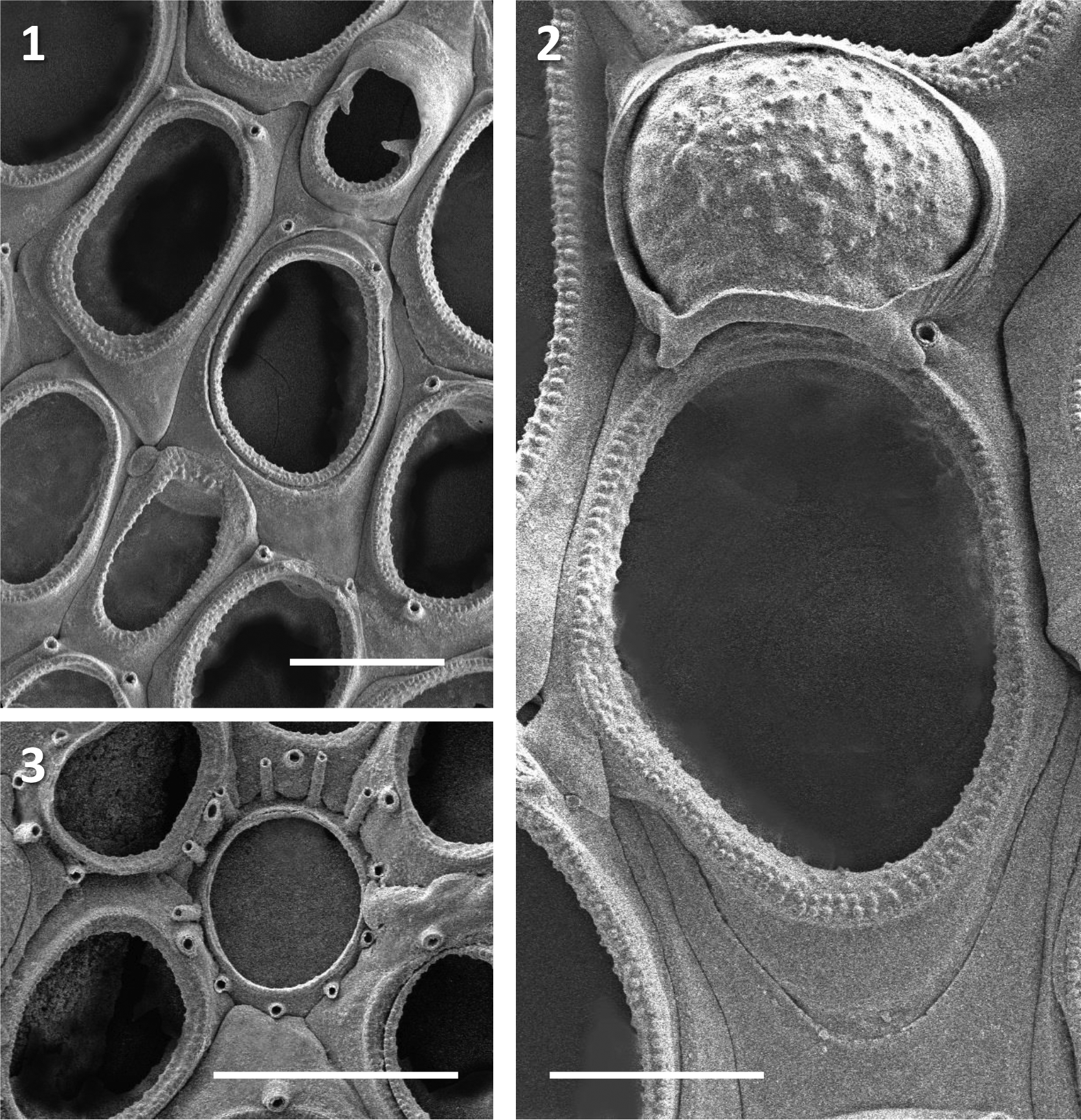
( Figs 1–3 View FIGURES 1 – 3 )
Membranipora kumatae Okada, 1923: 223 , figs 19, 20.
Crassimarginatella kumatae: Harmer 1926: 224 View in CoL ; Silén 1941: 24, fig. 19; Androsova 1958: 105, fig. 14; Mawatari & Mawatari 1980: 62, fig. 17 ( cum syn.); Hirose 2010: 31, pl. 50A, B.
Crassimarginatella crassimarginata: Rho & Seo 1986: 37 View in CoL , pl. 3; Seo 2005: 313, pls 33B, 34; Seo 2010: 26, fig. 10. Non Membranipora crassimarginata Hincks, 1880 .
Material examined. NIBRIV0000325934— RV Kuklipgongwon-yeoungu 1 stations I2, I3, J3, on small rocks and pebbles; Seogwipo , Jeju-do.
Description. Colony encrusting, unilaminar, up to 42 mm in diameter. Autozooids changing in proportion from squat and more or less suboval towards the colony center, longer and more elongate-oval in outer parts of colony; contiguous, arranged in regular quincunx, the boundaries between zooids marked by thin lines in shallow furrows. Gymnocyst smooth, moderately developed proximally, narrowing laterally and especially distally. Cryptocyst narrow, granular, of even width around the entire opesia. Periancestrular zooids with 4–5 slender spines, these rapidly diminishing in number to 2 on many zooids, with no spines on zooids in peripheral part of colony; some zooids with oral spines also have a medial spine on gymnocyst immediately proximal to cryptocyst. Avicularium subvicarious, less than half the size of an autozooid, surrounded by 4 autozooids; distal side elevated, cowl-like, proximal face steeply inclined, almost equally divided into rostral and opesial halves, separated by a robust pair of proximally angled acute condyles; proximal cryptocyst granular. Ooecium hyperstomial, ectooecium mostly membranous except for a narrow circumferential band of calcification; endooecium extensive, granular. Ancestrula tatiform, more or less circular, with large round opesia and 11 evenly spaced gymnocystal spines.
Measurements. ZL 265–402 (320) µm, ZW 152–308 (224) µm; OpL 189–271 (214) µm, OpW 137–179 (161) µm; OoL 136–156 (148) µm, OoW 152–197 (169) µm; AvL 183 µm, AvW 163 µm; AnL 142 µm, AnW 129 µm.
Remarks. Rho & Seo (1986) first recorded this species in Korean waters from Jeju Island, South Sea, as Crassimarginatella crassimarginata . This species, originally described from the Mediterranean Sea ( Hincks 1880), was also reported in Japanese waters by Mawatari & Mawatari (1980), although their material is neither conspecific with C. crassimarginata nor with that from Korea’s South Sea coasts. The species reported for South Korea is C. kumatae , first described from the Straits of Korea ( Okada 1923), whereas that of Mawatari & Mawatari (1980) may represent an undescribed species—it differs most notably from C. crassimarginata in having an ooecium with a membranous ectooecium and a large skeletal surface of granular endooecium. Harmelin (1973) illustrated C. crassimarginata from the Mediterranean Sea as having a skeletally smooth ectooecium with a narrow transverse slit frontally.
The marked difference in the frontal appearance of the ooecium in these taxa raises the question of the scope of the genus Crassimarginatella , as noted by Gordon (2014). On the one hand, the type species of the genus, C. crassimarginata , has as mentioned above a calcified ectooecium with a transverse slit. Some other species have a similar ooecium, in which there is either a narrow slit, e.g. Crassimarginatella papulifera ( MacGillivray, 1882) , or a broad furrow in which a smooth narrow exposure of endooecium can be seen, e.g. Crassimarginatella vincularia Gordon, 1984 . On the other hand, there is a cluster of species, predominantly in northeastern Asian waters but extending to New Zealand, which have a largely membranous ectooecium, with only a narrow peripheral band of calcification, and a large frontal exposure of granular endooecium, e.g. Crassimarginatella japonica ( Ortmann, 1890) , C. kumatae ( Okada, 1923) and Crassimarginatella spinifera Silén, 1941 from the northwestern Pacific, and Crassimarginatella cucullata ( Waters, 1898) from New Zealand. This latter group is cohesive morphologically although some characters vary—avicularia may have either a complete cross-bar or only pivots, and there may or may not be oral and opesial spines.
Could this cluster of species, which has every appearance of being a clade of genetically related taxa, be split off from Crassimarginatella sensu stricto? The problem is that of intermediate, or seemingly intermediate, morphological grades. For example, Crassimarginatella calva ( Brown, 1952) [inadvertently named and illustrated as Crassimarginatella electra Gordon, 1984 ] has an ooecium in which about half the ectooecium is calcified, leaving a fairly large area of endooecium, although this is smooth, not granular. Furthermore, like C. kumatae , this species has a widely separated pair of oral spines and a median gymnocystal spine. Evolutionarily, heterochrony could account for differences in expressions of the relative amounts of calcified ectooecium vs endooecium. Avicularian morphology seems to vary independently of ooecial morphology, inasmuch as vicarious and subvicarious forms, with or without complete cross-bars, are found in species with frontal ooecial fissures or large areas of granular endooecium. Our conclusion is that, until genetic information is available for a wide variety of Crassimarginatella species, it would be premature to split off a new genus that may render Crassimarginatella paraphyletic.
Distribution. Korea: South Sea coasts; Japan: Straits of Korea, Sagami Bay, Ogasawara (Bonin) Islands; 27– 300 m on various hard substrata.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Flustrina |
|
Family |
|
|
Genus |
Crassimarginatella kumatae ( Okada, 1923 )
| Min, Bum Sik, Seo, Ji Eun, Grischenko, Andrei V., Lee, Sang-Kyu & Gordon, Dennis P. 2017 |
Crassimarginatella crassimarginata :
| Seo 2010: 26 |
| Seo 2005: 313 |
| Rho 1986: 37 |
Crassimarginatella kumatae :
| Hirose 2010: 31 |
| Mawatari 1980: 62 |
| Androsova 1958: 105 |
| Silen 1941: 24 |
| Harmer 1926: 224 |
Membranipora kumatae
| Okada 1923: 223 |