
Rhinebothrium tetralobatum Brooks, 1977
|
publication ID |
https://doi.org/10.11646/zootaxa.4300.3.5 |
|
publication LSID |
lsid:zoobank.org:pub:EE5688F1-3235-486C-B981-CBABE462E8A2 |
|
DOI |
https://doi.org/10.5281/zenodo.6052031 |
|
persistent identifier |
https://treatment.plazi.org/id/03D087A8-FFB4-FFF3-ACE3-57A7FA24F851 |
|
treatment provided by |
Plazi |
|
scientific name |
Rhinebothrium tetralobatum Brooks, 1977 |
| status |
|
Rhinebothrium tetralobatum Brooks, 1977 , Family Rhinebothriidae Euzet, 1953 , order Rhinebothriidea
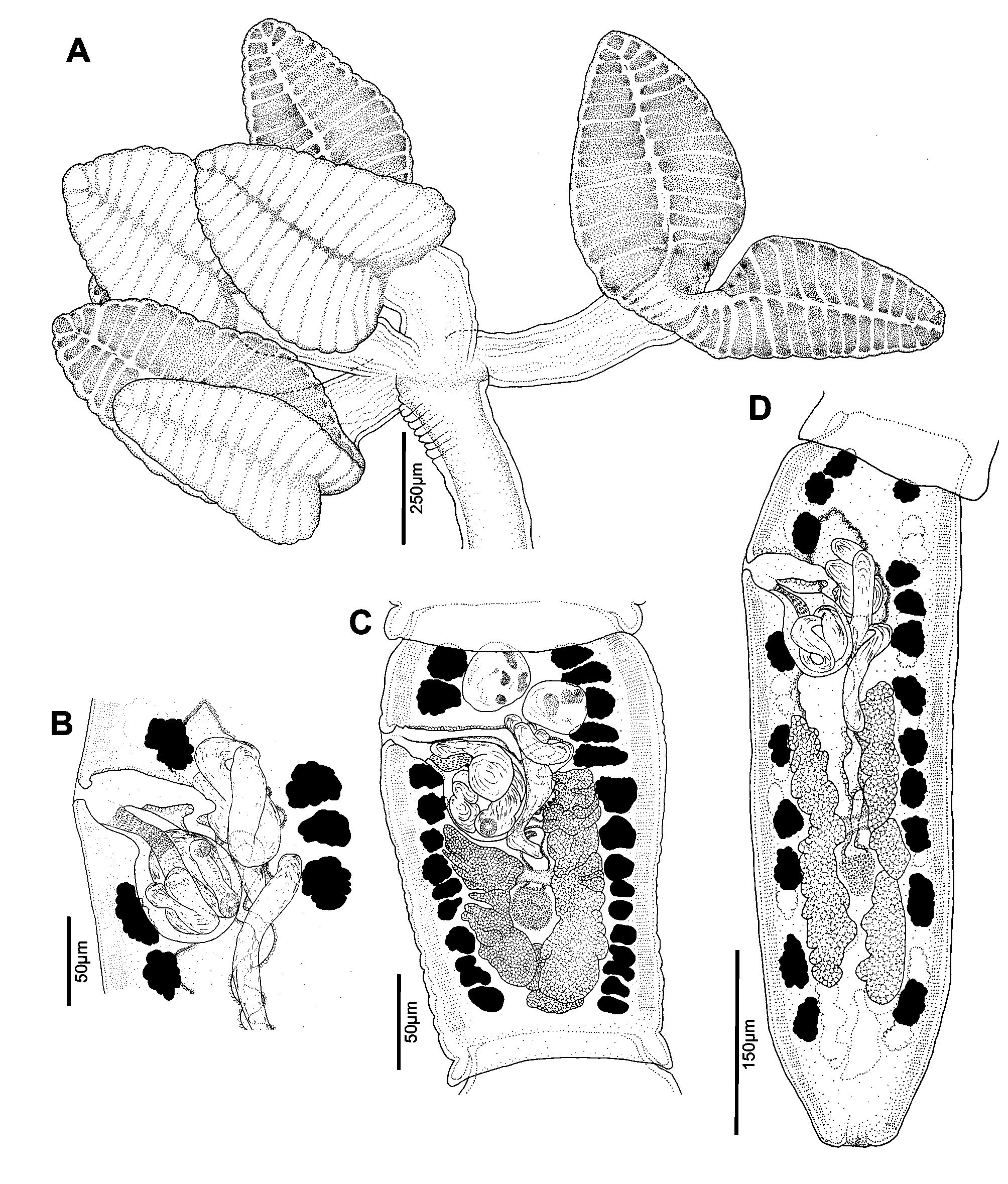
( Figs. 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 )
Type host: Styracura schmardae (Werner) de Carvalho, Loboda & da Silva ( Myliobatiformes : Potamotrygonidae ).
Type locality: Caribbean Sea , 15 km west of La Cienaga, Magdalena, Colombia ( 11°01'N, 74°15'W). GoogleMaps
Additional locality: Tasajeras , Magdalena, Colombia ( 10°58'N, 74°19'W and 11°0'N, 74°16'W). GoogleMaps
Site of infection: Spiral intestine.
Type material: Holotype (USNPC 73967) and 6 paratypes (HWML 20253, 20266).
Voucher specimens deposited: HWML 110067–110075 (9 complete worms); LRP 9254–9269 (14 complete worms); MZUSP 7926a–7926e and 7927a–7927d (9 complete worms), 7928–7929 (2 SEM vouchers), and 7930a– 7930c (voucher and histological sections).
Prevalence of infection and distribution ― 80% (four of five spiral intestines), seeming to be restricted to the Atlantic coast of Colombia.
Redescription. [Based on the type series, and 58 additional mature specimens, which includes 53 whole mounts, two worms observed with SEM, and three prepared as cross-sections]. Worms craspedote ( Fig. 1 View FIGURE 1 ), euapolytic, 4.1–19.0 mm (n = 42) long, composed of 80–206 (n = 42) proglottids. Scolex 278–801 (n = 48) in maximum width, composed of four stalked and bilobed bothridia constricted at center, with muscular rims ( Fig. 2 View FIGURE 2 A, 3A). Bothridia 487–1,094 (n = 48) long by 122–328 (n = 48) wide, divided by 23–34 (n = 48) transverse septa and one medial longitudinal septum into 47–69 (n = 32) loculi; with anterior-most loculus single, 26–50 (n = 31) long by 31–59 (n = 31) wide; posterior-most loculi double. Cephalic peduncle absent. Proximal and distal surface of bothridia covered with acicular filitriches and gladiate spinitriches ( Fig. 3 View FIGURE 3 B–G). Anterior portion of strobila covered with capilliform filitriches ( Fig. 3 View FIGURE 3 H).
Immature proglottids wider than long, becoming longer than wide with maturity, 68–178 (n = 42) in number ( Fig. 1 View FIGURE 1 ). Terminal mature proglottids 173–900 (n = 41) long by 73–220 (n = 42) wide ( Fig. 2 View FIGURE 2 C,D), 6–62 (n = 42) in number. Some terminal proglottids with sperm-filled vas deferens and atrophied testes ( Fig. 2 View FIGURE 2 D). Testes in anterior ¼ to ½ of proglottid, oval, 15–44 (n = 37) long by 10–32 (n = 37) wide, two (n = 38) in number, arranged opposite one another ( Fig. 2 View FIGURE 2 C, 4A). Cirrus sac in anterior ½ of proglottid, round to pyriform, 25–78 (n = 38) long by 28–69 (n = 38) wide, containing coiled cirrus; cirrus armed with spinitriches ( Fig. 2 View FIGURE 2 B). Genital atrium indistinct. Genital pore 18–33% (n = 41) of proglottid length from anterior end, irregularly alternating. Vagina opening into genital atrium anterior to cirrus sac, thick-walled, weakly sinuous, somewhat enlarged in proximal part, with antero-medial portion adjacent to cirrus sac, both directed laterally to common genital atrium. Vaginal sphincter absent. Ovary near posterior end of proglottid, bilobed in dorso-ventral view, tetralobed in cross-section ( Fig. 4 View FIGURE 4 B), symmetrical, 68–348 (n = 42) long by 40–112 (n = 42) wide at isthmus. Vitelline follicles extending length of proglottid, longer 7–24 (n = 15) than wide 4–19 (n = 15). Detached gravid proglottids and eggs not observed.
Remarks. The original description of Rhinebothrium tetralobatum was based on six specimens and did not include information on tegumental structures ( i.e., microtriches). In addition, we also detected some discrepancies between the original description and re-examination of the type series. Brooks (1977) reported R. tetralobatum as being 15–30 mm in length, with a genital pore position between 44 and 48% from anterior end of proglottid and an ovary measuring 198–300 in length. These accounts differ from our measurements of the type series ( i.e., 5.4–9.7 mm, 23–33%, and 86–159, respectively). Furthermore, the examination of additional material shows that R. tetralobatum ranges from 4.1 to 19.0 mm in length, possesses genital pores that are located 18 to 33% from the anterior end, and that the ovary is 68–348 in length ( Table 1). Also, Brooks (1977) reported the presence of a cephalic peduncle, which was not seen in either the newly collected specimens, or the type series, and described the worms as apolytic rather than euapolytic (sensu Caira et al. 1999).
Among 41 valid species of Rhinebothrium , R. tetralobatum shares the presence of two testes per proglottid with five other members of the genus. This group includes the freshwater species R. fulbrighti Reyda & Marques, 2011 ex Potamotrygon orbignyi Castelnau from the Bay of Marajó, Brazil; the marine Atlantic R. biorchidum Huber & Schmidt, 1985 ex Urobatis jamaicensis Cuvier from Jamaica; R. spinicephalum Campbell, 1970 ex Hypanus americanus (Hildebrand & Schroeder) from the coast of Virginia, U.S.A.; the eastern Pacific R. ditesticulum Appy & Dailey, 1977 ex Urobatis halleri Cooper from California, U.S.A.; and R. rhinobati Dailey & Carvajal, 1976 ex Rhinobatos planiceps Garman from the coast Antofagasta, Chile. Biogeographical distribution aside, R. biorchidum , R. rhinobati , and R. spinicephalum differ from R. tetralobatum by having fewer segments (15–26, 18–33, 36–49 vs. 80–206, respectively) and a smaller number of bothridial loculi (22–30, 22, 32–34 vs. 47–69, respectively). In addition, R. biorchidum and R. rhinobati can be further distinguished from R. tetralobatum by their smaller size (1.2–2.5, 1.8–2.8 mm vs. 4.1–19.0 mm, respectively).
Rhinebothrium ditesticulum and R. fulbrighti most closely resemble R. tetralobatum in their total length (9.6– 28.7 and 3.1–18.0 mm vs. 4.1–19.0, respectively) and the number of proglottids (160–276 and 40–168 vs. 80–206, respectively). Besides the slightly overlap, R. fulbrighti possesses fewer bothridial loculi in comparison to R. tetralobatum (43–53 vs. 47–69, respectively). In addition to that, the former has been described as having only two ovarian lobes whereas the latter has four. Rhinebothrium ditesticulum can be distinguished from R. tetralobatum by the morphology of the bothridia. It has a single posterior-most loculus on the bothridia and the anterior and posterior bothridial surfaces are completely separated, whereas, R. tetralobatum possess two posterior-most loculi and the anterior and posterior bothridial surfaces are not completely separated, but have a constriction at the center.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Eucestoda |
|
Order |
|
|
Family |
|
|
Genus |