
Rhinebothrium reydai, Trevisan & Marques, 2017
|
publication ID |
https://doi.org/10.11646/zootaxa.4300.3.5 |
|
publication LSID |
lsid:zoobank.org:pub:EE5688F1-3235-486C-B981-CBABE462E8A2 |
|
DOI |
https://doi.org/10.5281/zenodo.6052033 |
|
persistent identifier |
https://treatment.plazi.org/id/7CF130AA-DCE7-4C07-9583-2415C96D2708 |
|
taxon LSID |
lsid:zoobank.org:act:7CF130AA-DCE7-4C07-9583-2415C96D2708 |
|
treatment provided by |
Plazi |
|
scientific name |
Rhinebothrium reydai |
| status |
sp. nov. |
Rhinebothrium reydai n. sp., Family Rhinebothriidae , order Rhinebothriidea
( Figs. 5–8 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 )
Type host: Styracura schmardae (Werner) de Carvalho, Loboda & da Silva ( Myliobatiformes : Potamotrygonidae ).
Type locality: Caribbean Sea off Almirante, Bocas Del Toro Province, Panama ( 09°17'N, 82°20'W and 09°17'N, 82°21'W). GoogleMaps
Site of infection: Spiral intestine.
Type specimens: MIUP CR1 (Holotype) and CRP1–CRP9 (Paratypes: 9 complete worms); MZUSP 7931a– 7931p (Paratypes: 16 complete worms), 7932 (1 SEM vouchers), and 7933a–7933d (voucher and histological sections); HWML 110076–110082 (Paratypes: 7 complete worms); LRP 9270–9276 (Paratypes: 7 complete worms).
Prevalence of infection and distribution: 45,5% (five of 11 spiral intestines), seeming to be restricted to the Caribbean coast of Panama.
Etymology: The species is named in honor of Dr. Florian B. Reyda, who took part in the sampling trip to Panama and who has contributed to the knowledge of this group of cestodes for many years.
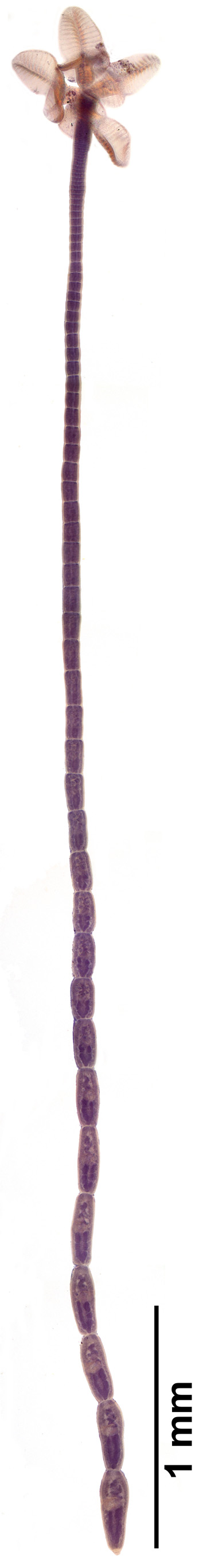
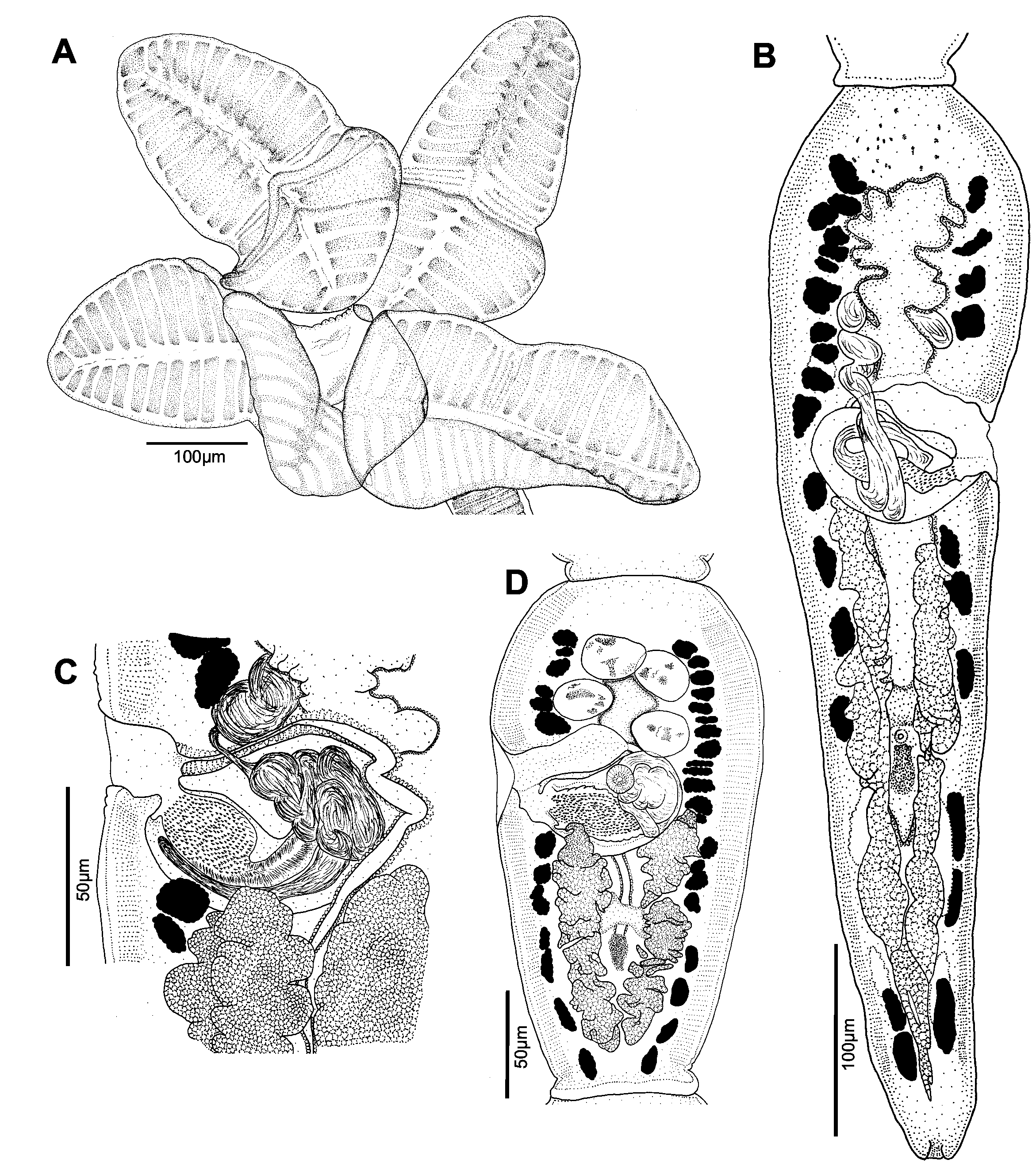
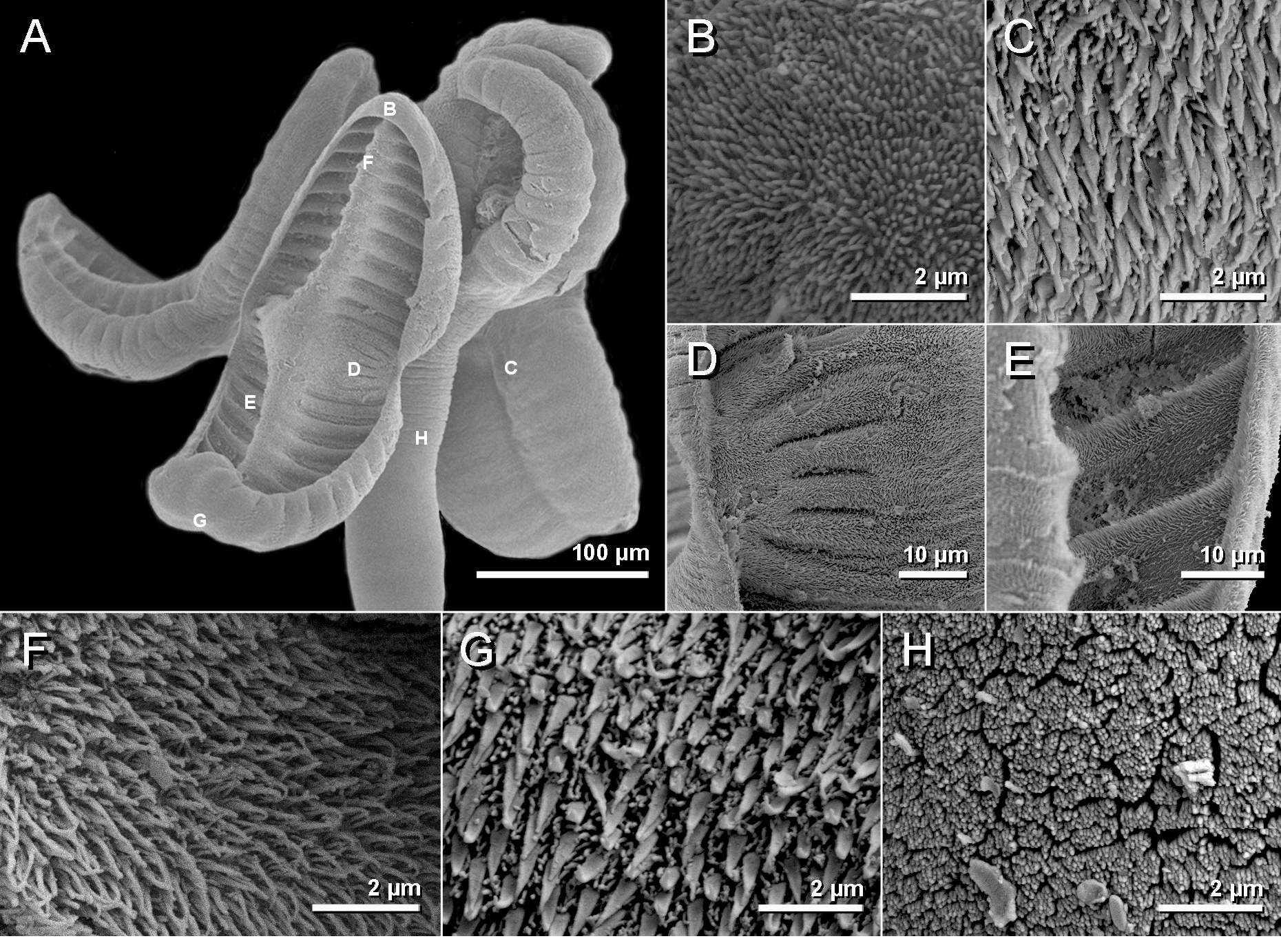
Description. [Based on 50 mature specimens: 47 whole mounts, one worm observed with SEM, and two prepared as cross-sections]. Worms acraspedote ( Fig. 5 View FIGURE 5 ), euapolytic, 2.2–7.4 mm (n = 47) long, composed of 26–68 (n = 47) proglottids. Scolex 237–376 (n = 8) in maximum width composed of four stalked and billobed bothridia constricted at center, with muscular rims ( Fig. 6 View FIGURE 6 A, 7A). Bothridia 280–461 (n = 9) long by 111–202 (n = 9) wide, divided by 17–22 (n = 15) transverse septa and one medial longitudinal septum into 34–44 (n = 15) loculi; anteriormost and posterior-most loculus single. Anterior-most loculus 18–26 (n = 10) long by 29–41 (n = 10) wide. Cephalic peduncle absent. Proximal and distal surface of bothridia covered with acicular filitriches and gladiate spinitriches ( Fig. 7 View FIGURE 7 C–G), except for anterior loculus with acicular filitriches only ( Fig. 7 View FIGURE 7 B). Anterior portion of strobila covered with capilliform filitriches ( Fig. 7 View FIGURE 7 H).
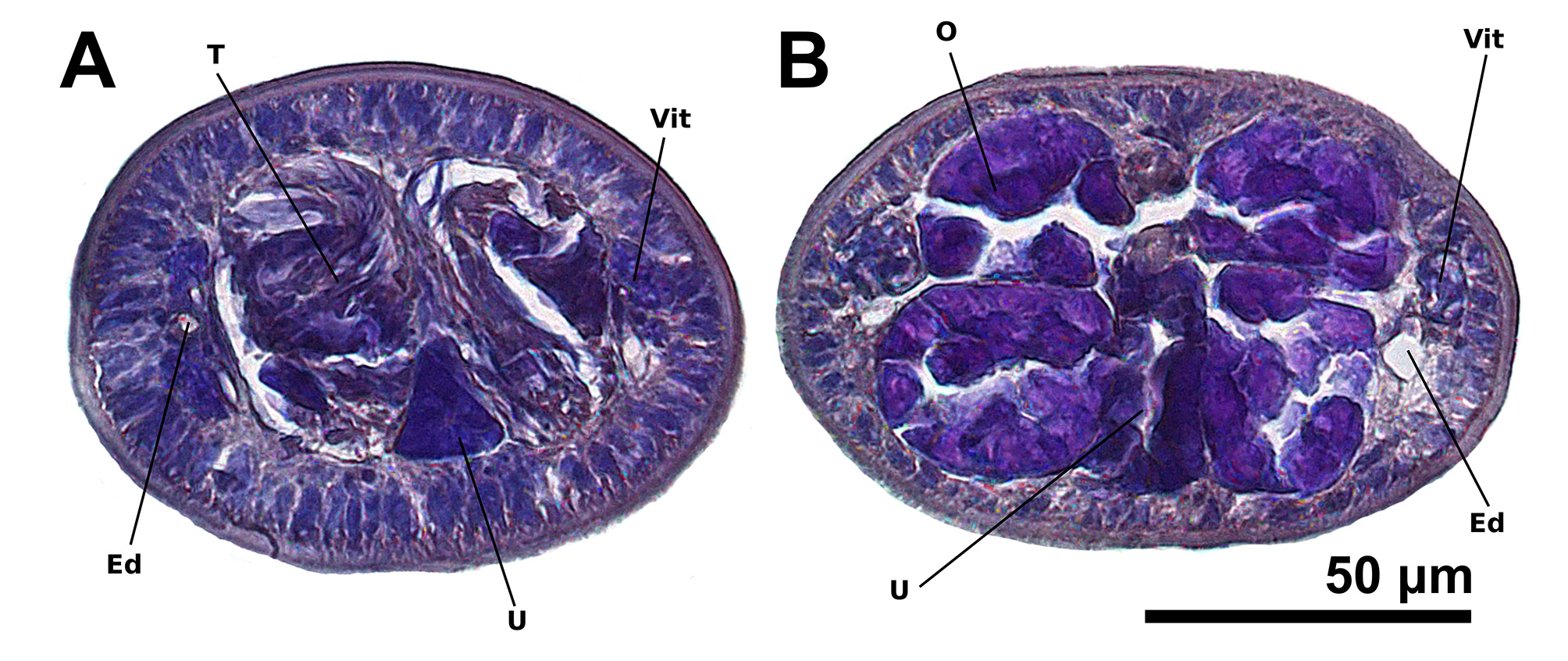
Immature proglottids wider than long, becoming as long as wide within the anterior third of the strobila (13– 30%, n = 39) thereafter, longer than wide with maturity, 19–56 (n = 47) in number ( Fig. 5 View FIGURE 5 ). Terminal mature proglottids ( Fig. 6 View FIGURE 6 B, D) 218–554 (n = 42) long by 67–146 (n = 43) wide, 4–14 (n = 47) in number. Some terminal proglottids with sperm-filled vas deferens and atrophied testes ( Fig. 6 View FIGURE 6 B). Testes in two irregular columns, in anterior ¾ of proglottid, oval, 20–43 (n = 27) long by 17–34 (n = 27) wide, 4 (n = 47) in number ( Fig. 6 View FIGURE 6 D, 8A). Cirrus sac in anterior ½ of proglottid, spherical to pyriform in shape, 33–59 (n = 39) long by 37–83 (n = 39) wide, containing coiled cirrus; cirrus armed with spinitriches ( Fig. 6 View FIGURE 6 C). Genital atrium prominent. Genital pores 31–49% (n = 42) of proglottid length from anterior end, irregularly alternating. Vagina opening into genital atrium anterior to cirrus sac, thick-walled, weakly sinuous, somewhat enlarged in proximal part, with antero-medial portion adjacent to cirrus sac, both directed laterally to common genital atrium. Vaginal sphincter absent. Ovary near posterior end of proglottid, inverted A-shaped in frontal view and tetra-lobed in cross-section ( Fig. 8 View FIGURE 8 B), symmetrical, 89–316 (n = 40) long by 39–79 (n = 40) wide at isthmus ( Fig. 6 View FIGURE 6 B). Vitelline follicles extending length of proglottid, longer 5–15 (n = 14) than wide 4–12 (n = 14). Detached gravid proglottids and eggs not observed.
Remarks. This new taxon is unique among species of Rhinebothrium due to a combination of morphological characters, including the presence of only four testes and a single posterior-most bothridial loculus. Compared to other 11 species that share the presence of four testes, only R. chollaensis Friggens & Duszynski, 2005 possesses a single posterior-most bothridial loculus. Rhinebothrium reydai n. sp. further resembles R. chollaensis in total length (2.2–8.4 vs. 1.3–5.1, respectively), number of proglottids (26–68 vs. 32–84, respectively), and number of bothridial loculi (34–44 vs. 40–49, respectively). Both species also have similar bothridial morphology. Friggens & Duszynskii (2005) described R. chollaensis as having leaf-like bothridia with a slight constriction at the level of the stalk, which is similar to what we observed in R. reydai n. sp.. However, after examining images from the type series of R. chollaensis ( Holotype [USNM 92213] and five paratypes [USNM 92213a–92214d]), we think that the constriction is more pronounced in the former, hence described as bilobed. Despite a similar morphology, R. chollaensis is apolytic whereas R. reydai n. sp. is euapolytic. Based on the type series, both species differ on the morphology of the aporal lobe of the ovary, which reaches the mid-lateral margin of the cirrus sac in R. chollaensis , whereas in R. reydai n. sp. the aporal lobe only reaches the posterior margin, since the cirrus sac takes approximately ¾ of the proglottid in width. Also, in R. reydai n. sp. the first square proglottid occurs within the anterior third of the strobila (13–30%, n = 39), whereas in the type series of R. chollaensis it occurs near the middle of the strobila (42–62%, n = 6). The morphological resemblance between these two species and their biogeographical distributions ( i.e., R. chollaensis from the eastern Pacific Ocean vs. R. reydai n. sp. from the western Atlantic Ocean), might also suggest a close phylogenetic association, which has yet to be tested within the context of a phylogenetic study.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Eucestoda |
|
Order |
|
|
Family |
|
|
Genus |