
Goniglossum liat, Freidberg, Amnon, 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4144.1.2 |
|
publication LSID |
lsid:zoobank.org:pub:F1A6E9DE-2CF3-4C4E-8E7F-29F44476CE15 |
|
DOI |
https://doi.org/10.5281/zenodo.6086599 |
|
persistent identifier |
https://treatment.plazi.org/id/03D087C8-4317-9D30-5FB3-FBF5FCB9F89C |
|
treatment provided by |
Plazi |
|
scientific name |
Goniglossum liat |
| status |
sp. nov. |
Goniglossum liat , n. sp.
Figs. 2 View FIGURE 2 , 3, 4 View FIGURES 3 – 5 , 6, 8–11 View FIGURES 6 – 8 View FIGURES 9 – 12 , 13, 15–24 View FIGURES 13 – 15 View FIGURE 16 View FIGURES 17 – 18 View FIGURES 19 – 22 View FIGURE 23 View FIGURE 24
Goniglossum wiedemanni, Freidberg and Kugler, 1989 [misidentification]. Carpomya wiedemanni, Norrbom, 1997: 340 View in CoL [ Israel record–misidentification]; Norrbom et al. 1999: 115 [ Israel record– misidentification].
Diagnosis. A detailed diagnosis and comparison with the only other congener is given above in the key to species of Goniglossum .
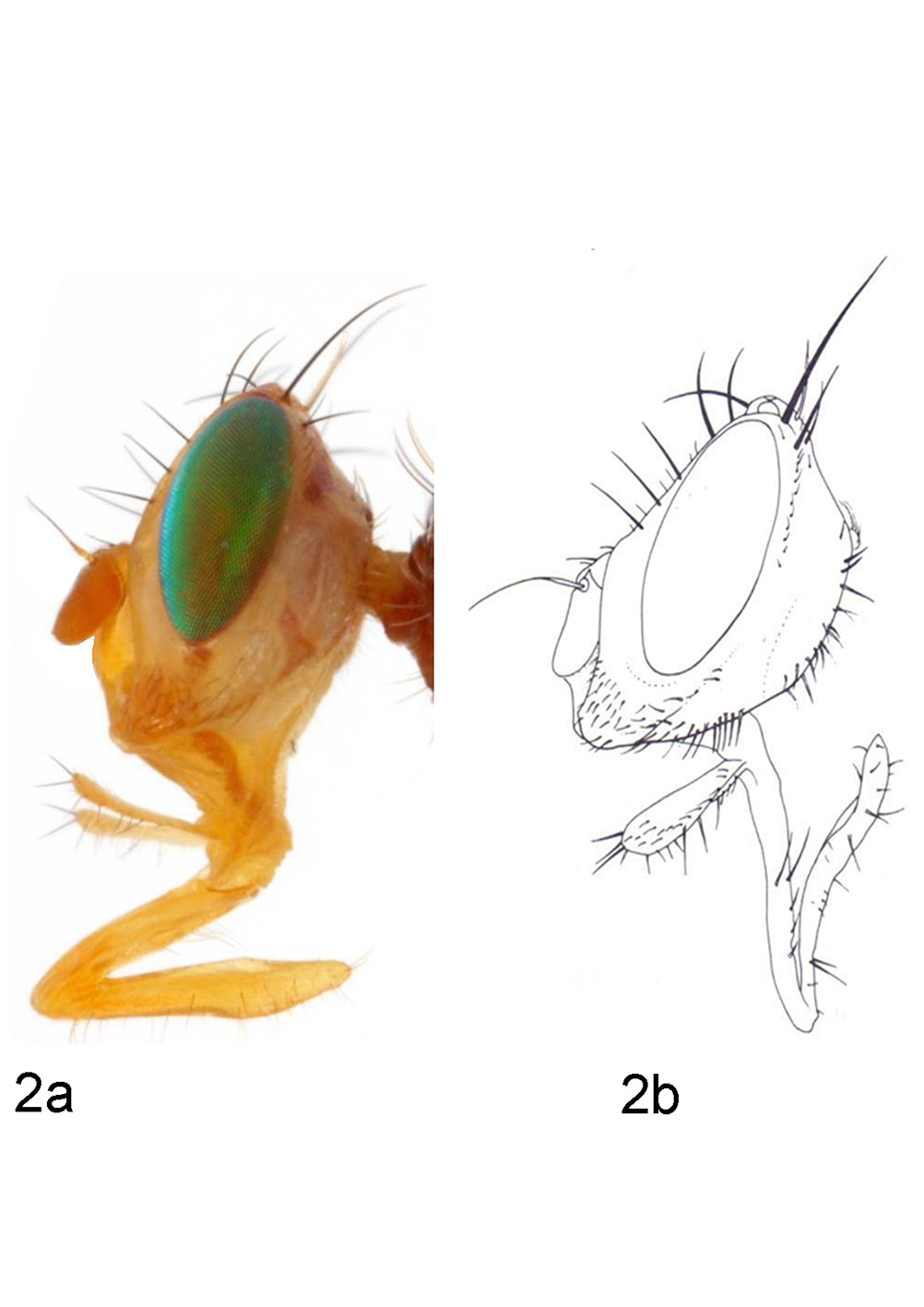
Description. Head ( Fig. 2 View FIGURE 2 ): Yellow; frons, face and gena mostly dull; occiput, ocellar plate and vertical plate shiny; parafacial and gena with fine white microtrichia, sometimes without sharp transition zone between dull and shiny areas; setulae mostly brown, on occiput and frontal plate yellowish; length:height:width ratio = 8.1:8.2:10.0; frons about as wide as long, at vertex 2.25 times as wide as eye, slightly narrower anteriorly; eye 2.3 times as high as long; parafacial as wide as antennal height; gena about 1.5 times as wide as antennal height.
Thorax ( Figs. 3, 4 View FIGURES 3 – 5 , 6 View FIGURES 6 – 8 ): Scutal pattern strongly variegated; mesonotal pattern comprises wide, black, usually complete median vitta, sometimes somewhat paler anteriorly, narrowed at or near base of acrostichal seta, broadened again, and extending slightly onto scutellum; dark pattern branching twice: first anterior to transverse suture, extending onto notopleuron, slightly onto postpronotal lobe and across transverse suture; secondly at median end of transverse suture, branch extending posteriorly as black stripe, then laterally towards pleuron, to or nearly to base of wing; black spot present on posterolateral corner of scutum, extending onto basal corner of scutellum; apex of scutellum with single quadrangular black spot, extending over bases of apical setae, rarely interrupted by small yellow spot centrally; remaining part of mesonotum yellow; subscutellum and mediotergite shiny black; 2 elongate areas of dense gray microtrichia present over anterior part of black median mesonotal vitta, microtrichia sometimes present also on black lateral branches of pattern; pleural pattern with 2 yellow to black wide longitudinal vittae (or areas) over an always distinctly and contrastingly paler, ivory-white background; dorsal vitta extends from anterior spiracle and propleuron over ventral part of anepisternum and anepimeron to base of halter, although this vitta much paler at anterior half and often black or blackish on posterior half; ventral vitta or area (mostly triangular) extends over ventral part of katepisternum and katepimeron. Setulae yellowish, rather coarse, dense on anterior and median part of mesonotum, sparser on pleuron, almost lacking from margin of scutum and scutellum.
Wing ( Fig. 8 View FIGURES 6 – 8 ): The extent of the dark pattern and the amount and extent of its various components (gray, yellow and brown) somewhat vary, but are not significantly different from those of the congener. Calypteres white; halter basally yellow, knob yellow or brownish.
Abdomen: Shiny yellow or reddish-yellow, with pairs of black spots anteriorly on tergites 2–5 in male, 2–5 or 2–6 in female; these pairs of spots sometimes united medially to form complete transverse bands; spots sometimes indistinct or entirely lacking, or anterior part of syntergite 1+2 with pair of small such spots laterally. Abdomen without obvious microtrichia. Tergite 6 of female 0.50–0.66 times as long as tergite 5. Setae at posterior margin of posteriormost 2 tergites black. Setulae on abdomen and oviscape yellowish to black.
Male terminalia ( Figs. 9–11 View FIGURES 9 – 12 ): Epandrium ( Figs. 9, 10 View FIGURES 9 – 12 ) as for genus. Phallus ( Fig. 11 View FIGURES 9 – 12 ) as for genus, bubble-like structure large, about 1.5 times as long as part of sclerotization distal to bubble.
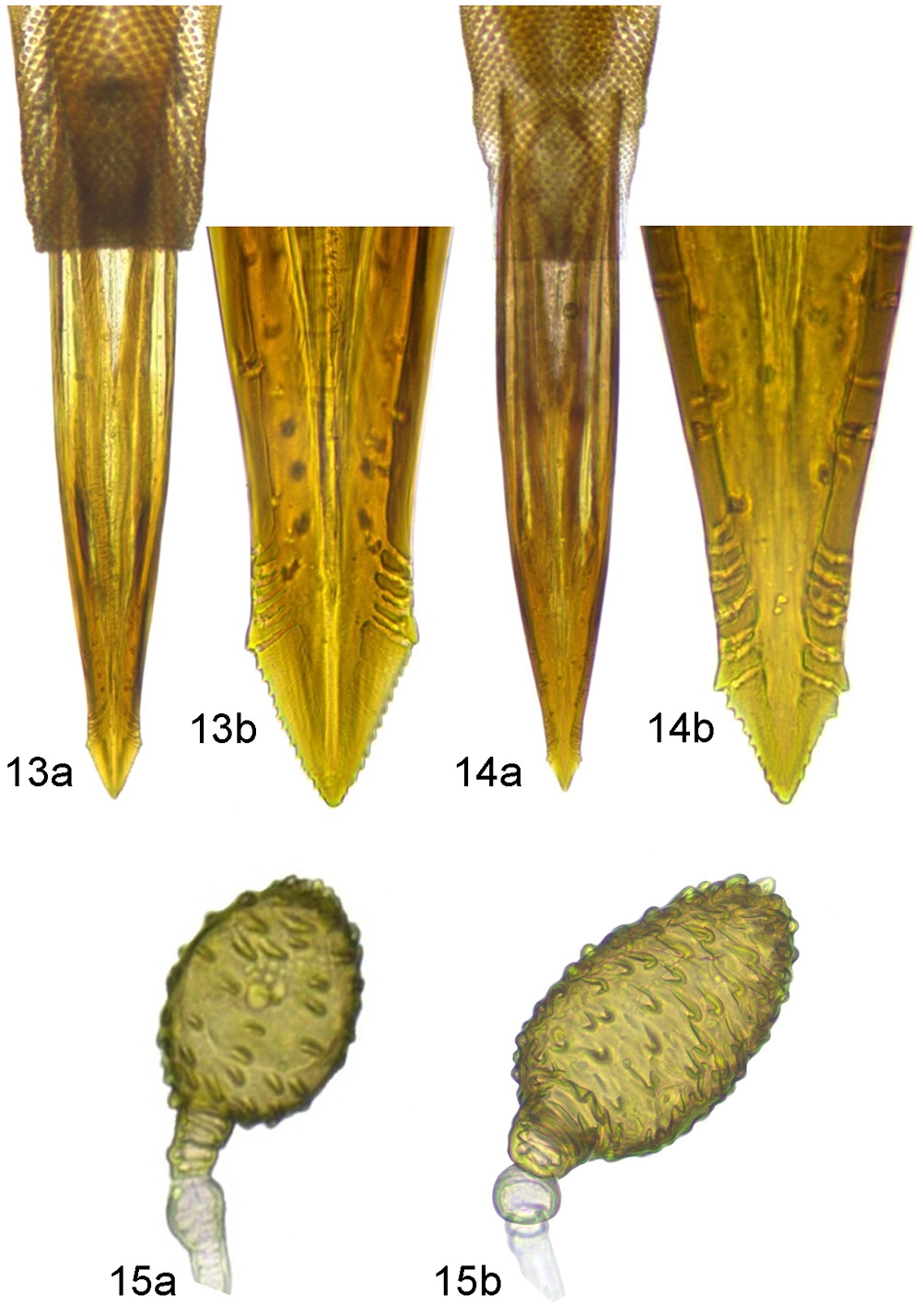
Female terminalia ( Figs. 13, 15 View FIGURES 13 – 15 ): Oviscape slightly longer than tergite 6, yellowish, with black apex. Aculeus ( Fig. 13 View FIGURES 13 – 15 a): aculeus-width-ratio averages 2.14 (range: 1.91–2.37); aculeus-shoulders-ratio averages 2.02 (range: 1.84–2.21); serration of arrow-head apex averaging 13.3 teeth (range: 11–15; Fig.13 View FIGURES 13 – 15 b). Spermathecae ( Fig. 15 View FIGURES 13 – 15 ) mostly spherical or ovoid (up to about 1.5 times as wide as long; Fig. 15 View FIGURES 13 – 15 a), sometimes elongate (up to about twice as long as wide; Fig. 15 View FIGURES 13 – 15 b).
Measurements (length in mm): Male: Body: 3.3–5.1, wing: 2.8–3.8. Female: Body: 4.5–5.5, wing: 3.6–4.5, oviscape: 0.25–0.30 (dorsal), 0.5–0.7 (ventral).
Material examined. [coordinates in brackets added while preparing this section; otherwise brackets indicate information not printed on the labels]: Holotype ♀, ISRAEL: [Southern Coastal Plain:] Nizzanim , [31°44.5'N 034°37'E], [reared from fruit collected in] iii–iv.2005, A. Freidberg, ex [fruit] Bryonia syriaca , [adult emerged on:] 1.iii–12.iv.2006, #14379 (SMNHTAU) GoogleMaps . Paratypes (almost all in SMNHTAU); same collection data as holotype (64♂, 69♀); same locality: iii–iv.[20]05, A. Freidberg, ex fruit Bryonia syriaca , [adult:] 26.ii–5.iii.2006, #14379 (60♂, 40♀) GoogleMaps ; same collection data, but not reared (4♂, 14♀). Additional paratypes (localities arranged from north to south and from west to east): ISRAEL: Har Hermon , 1600m, [33°18'N 035°46'E], 5.viii.2004, A. Freidberg and L. Lerner, ex fruit Bryonia cretica , pupa GoogleMaps : 23.viii.–2.ix.2004; adult: 15.v.2005 (23♂, 22♀); same collection data, but slightly different dates, L. Lerner (8♂, 1♀); same collection data except adult: 20–30.iv.2008 (8♂, 4♀). Har Hermon , 1650m, 17.viii.2005, A. Freidberg, ex Bryonia cretica fruit, pupa : 17.viii–10.ix.05, adult: 1–5.v.2006 (10♂, 8♀); adult: 25.iv.2008 (20♂, 8♀); Senir [33°14.5'N 35°40'E], summer 1995, A. Freidberg, ex berry Bryonia cretica , v.1996 (24♂, 21♀) GoogleMaps ; Panyas , 33°15'N 35°42'E, 8.v.2010, A. Freidberg (1♀) GoogleMaps ; ’ Alma [33°03'N 35°30'E], 10.vi.1992, A. Freidberg, ex Bryonia cretica berry, pupa GoogleMaps : 11.vi–6.vii.1992, adult: iv.1994, #9735 (18♂, 15♀); ’ Alma , summer 1995, A. Freidberg, ex berry Bryonia cretica , v.1996 (24♂, 21♀) ; Har [Mt.] Meron , 800m, 13.vii.1971, [32°59.5'N 035°24'E], A. Freidberg, ex Bryonia cretica [written multiflora], 20.iv.1973 GoogleMaps – 10.v.1973, #9661 (5♂, 6♀); Har Meron , 1100m, 22.v.1994, 31°52'N 35°24'E, A. F reidberg and F. Kaplan, #9749 (1♀) GoogleMaps ; Kefar Shammay , 2.v.1994, [32°57.3'N 35°27.2'E], A. Freidberg, #9741 (2♂) GoogleMaps ; Park haYarden [32°55'N 35°33'E], 14.iv.1999, A. Freidberg, #9348 (1♂) GoogleMaps ; Kare Deshe , v–vi.1973, [32°52.3'N 35°32.5'E], A. Freidberg, ex Bryonia cretica [written multiflora] iii–iv.1974 (2 ♂) GoogleMaps ; 15.iv.1992, A. Freidberg, #9686 (8♂, 6♀); Hof Rotem-Shezaf, 3kmS ' En Gev , - 200m, 32°46'N 35°38.3'E, 21.iii.2010, A. Freidberg (2♂) GoogleMaps ; E. Morgulis (1♂); Carmel , iv.1983, [32°47.5'N 34°58.5'E], A. Freidberg, ex Bryonia , pupa GoogleMaps : v.1983, adult: iv.1985, #9565 (3♀); Nahal Oren Spill , 30.iv.2008, A. Freidberg, ex fruit Bryonia cretica , puparium: v.[20]08, adult : 20.iv. [20]09 (1♂, 3♀); Qiryat Tiv'on , 7.iii.1981, [32°43'N 35°08'E], A. Freidberg, #9685 (5♂, 1♀) GoogleMaps ; Ginnegar , 15.iv.1981, [32°39.5'N 35°15'E], T. Furman, adult GoogleMaps : 15–17.iii.1982, #9681 (15♂, 16♀); Umm el-Ghanem, 8.v.2007, [32°40'N 35°24'E], A. Freidberg (2♀) GoogleMaps ; Nahal Tavor, North facing slope [32°37'N 35°32'E], 26.iii.2001, L. Friedman, #9349 (1♀) GoogleMaps ; Ramat Dawid [32°40.5'N 35°12'E], 28.iii.1981, T. Furman, #9684 (1♂, 1♀) GoogleMaps ; ' En haNaziv [32°28.2'N 35°30'E], 18.iv.1996, A. Freidberg, # 9746 (2♂, 3♀) GoogleMaps ; Hadera [32°25'N 34°54'E], 26.vii.2006, A. Freidberg, ex Bryonia cretica , larva GoogleMaps : 6–22.viii.2006, adult: 10–15.iv.2007 (32♂, 30♀); 21.x.2003, L. Lerner, ex fruit Bryonia cretica , pupa: 23–31.x.2003, adult: 17.iii.2005 (2♂, 3♀); 24.iv.2004, A. Freidberg (2♂); 28.iv.2004, L. Lerner (4♂, 5♀); Berekhat Ya'ar [32°24'N 34°54'E], 28.iv.2004, A. Freidberg (1♂, 3♀) GoogleMaps ; Yarhiv , 24.iii.1983, [32°09'N 34°58'E], A. Freidberg (1♂, 2♀) GoogleMaps ; Kokhav Ya'ir, Ya'ar Sappir , 160m, 32°13.9'N 34°59.5'E, 16.ii.2010, L. Friedman (1♀) GoogleMaps ; 32°14'N 34°59'E, 21.iii.2011, L. Bodner (2♂, 1♀); A. Freidberg (2♀); Herzliyya [32°10'N 34°50'E], 1.vi.2008, ex fruit Bryonia cretica , puparium: 5–25.vi. [20]08, adult: 10–15.iv. [20]09 (16♂, 22♀) GoogleMaps ; Zomet Gelilot , [32°09'N 34°49'E], 3.viii.2006, A. Freidberg, ex fruit Bryonia cretica , pupa: 22.vii–ix. [2007], adult GoogleMaps : 10–15.iv.2007 (12 ♂, 13♀); Zomet Gelilot , orchard, shade, 15.ix.–30.x.2007, A. Freidberg, ex [fruit] Bryonia cretica , adult : 5.iv.2008 (6♂, 3♀); Tel Aviv, Gelilot , 32°9.33'N 34°48.46'E, 19.iii.2005, A. Freidberg (10♂, 5♀) GoogleMaps ; Tel Aviv, 1.iii.1973, [32°08'N 34°49'E], A. Freidberg, #9670 (1♂) GoogleMaps ; Latrun , 10.iii.1973, [31°39.5'N 34°58.5'E], A. Freidberg, #9566 (2♂, 1♀) GoogleMaps ; Bet Shemesh , 31°44'N 34°58'E, 30.iii.2004, L. Lerner, ex Bryonia fruit, 5.iii.2005 (6♂, 6♀) GoogleMaps ; Bet ' Oved [31°55.5'N 34°46'E], 27.iv.2004, A. Freidberg (1♂) GoogleMaps ; Yavne , 18.ii.1971, [31°52'N 34°44.7'E], A. Freidberg, #9560 (5♂, 4♀), 24.ii.1971, A. Freidberg ,, #9562 (2♂, 3♀), 1.iv.1971, A. Freidberg, ex Bryonia syriaca , 23.ii.1973, #9673 (3♂, 1♀), 26.ii.1972, A. Freidberg, #9571 (7♂), 11.iii.1972, A. Freidberg, #9561 (4♂, 2♀) GoogleMaps ; Nizzanim , [31°44.5'N 034°37'E], iii–iv.2005, A. Freidberg, ex [fruit] Bryonia syriaca , 1.iii–12.iv.2006, #14379 (64♂, 70♀) GoogleMaps ; Nizzanim , iii–iv.[20]05, A. Freidberg, ex fruit Bryonia syriaca , 26.ii–5.iii.2006, #14379 (60♂, 40♀) ; same collection data, but not reared (4♂, 14♀); Ashqelon , 2.iii.1974, [31°40'N 34°34'E], A. Freidberg, #9675 (1♂, 1♀) GoogleMaps ; Bet Guvrin , 31.iii.1975, [31°36.7'N 034°53.7'E], A. Freidberg, #9677 (1♀) GoogleMaps ; Sederot , 27.ii.1974, [31°32'N 34°35.3'E] A. Freidberg, #9676 (1♂, 1♀) GoogleMaps . Other material (not paratypes): TURKEY: Termessos , Antalya, 20KmNW, 800– 1000m, [36°59'N 30°28'E], 3.V.2000, A. Freidberg & H. Ackerman (2♂) GoogleMaps .
The holotype is double-mounted (minutien pin on a plastic block), is in excellent condition and is deposited in SMNHTAU. Most of the paratypes are deposited in SMNHTAU, but many have been distributed among numerous museums and collections (see list of collection acronyms in the Material and Methods section).
Biology and host plants. My students and I acquired most of the information provided in this section over the course of many years of field collecting, and through observations in both the field and laboratory (Freidberg, personal data). Almost all my observations and conclusions were substantiated methodically by my M.Sc student, Mrs Larisa Lerner-Alfi (2007), during her study, for which I am grateful. The most interesting results concerning the new species are summarized briefly in the next few paragraphs.
In the laboratory, copulation begins with the male mounting the female, apparently without any obvious courtship. Mating position is similar to positions described for many other tephritids (e.g., Freidberg, 1982). Crossmating between G. liat and G. wiedemanni ( Fig. 16 View FIGURE 16 ) observed during the hybridization experiments mentioned above was similar to normal single-species mating observed in G. l i a t. Both male and female are promiscuous, mating multiple times over the course of their lifetime, often almost every day, with the same or different mates. Copulation, based on laboratory observations, is unusually long, averaging 7.50 hours (range: 0.30–20.00 hours), and begins around noon, when many, often most, flies in a cage were observed to mate simultaneously.
Both Bryonia species in Israel, i.e., B. cretica L., and Bryonia syriaca Boiss. (Cucurbitaceae) , are climber plants ( Fig. 17 View FIGURES 17 – 18 ), with the former species sometimes climbing to heights of 10 m and more on various trees. B. cretica is dioecious, with bunches of numerous female flowers ( Fig. 18 View FIGURES 17 – 18 ) and larger male flowers. Young fruits are the usual substrate for oviposition by these fruit flies, and oviposition ( Fig. 19 View FIGURES 19 – 22 ) is often immediately followed by a behavior which we speculate to be host-marking, and which probably prevents repeated oviposition by conspecifics into the same fruit. This behavior comprises moving straight ahead while dragging the tip of the extended aculeus ( Fig. 20 View FIGURES 19 – 22 ) and swinging it from side to side in a zigzag manner. Every few seconds, a sharp change in the direction of movement takes place.
In nature, females use their ovipositor not only to deposit eggs and for host-marking, but also to drill into green fruits and then suck at the sap ( Fig. 19 View FIGURES 19 – 22 , arrow) that oozes from the wound. Another possible source of nourishment for the adult flies is the Bryonia flower, and more precisely its nectar content. I have often observed individuals of G. liat inserting their head and proboscis into Bryonia flowers, apparently to feed on the nectar ( Fig. 21 View FIGURES 19 – 22 ). These flies were often also observed on male flowers ( Fig. 22 View FIGURES 19 – 22 ) with their body covered in pollen. Although the flies are not assumed to feed on this pollen they may well serve as pollinators for these host plants.
The larvae develop in the berries of Bryonia cretica L. and B. syriaca Boiss. Each infested fruit generally yielded a single larva that left this fruit prior to pupation, although during a massive fruit dissection we recorded several cases in which two individuals shared the same fruit, albeit one of them would invariably die (we indeed observed some dead larvae in dissected fruits that also contained an healthy larva).
Fruits collected on Mt. Hermon (see Material Examined) in summer 2004 contained larvae that pupated during the next few months. Adults from the same lot of pupae emerged in the next spring (2005), but also in the spring of 2008, demostrating that adult flies can emerge up to at least four years after pupation. This is a phenological phenomenon that, although possibly more common than expected, has seldom been reported in the literature ( Bateman, 1972).
A somewhat similar phenomenon was detected while studying the egg+larva stage of the phenology. Samples of thousands of fruits were transferred to the lab almost monthly for one year. We were surprised to discover that larvae were still emerging from the fruits up to half a year following egg deposition. We termed this developmental stage “egg+larva”, due to the inability to time the hatching of the first instar larva.
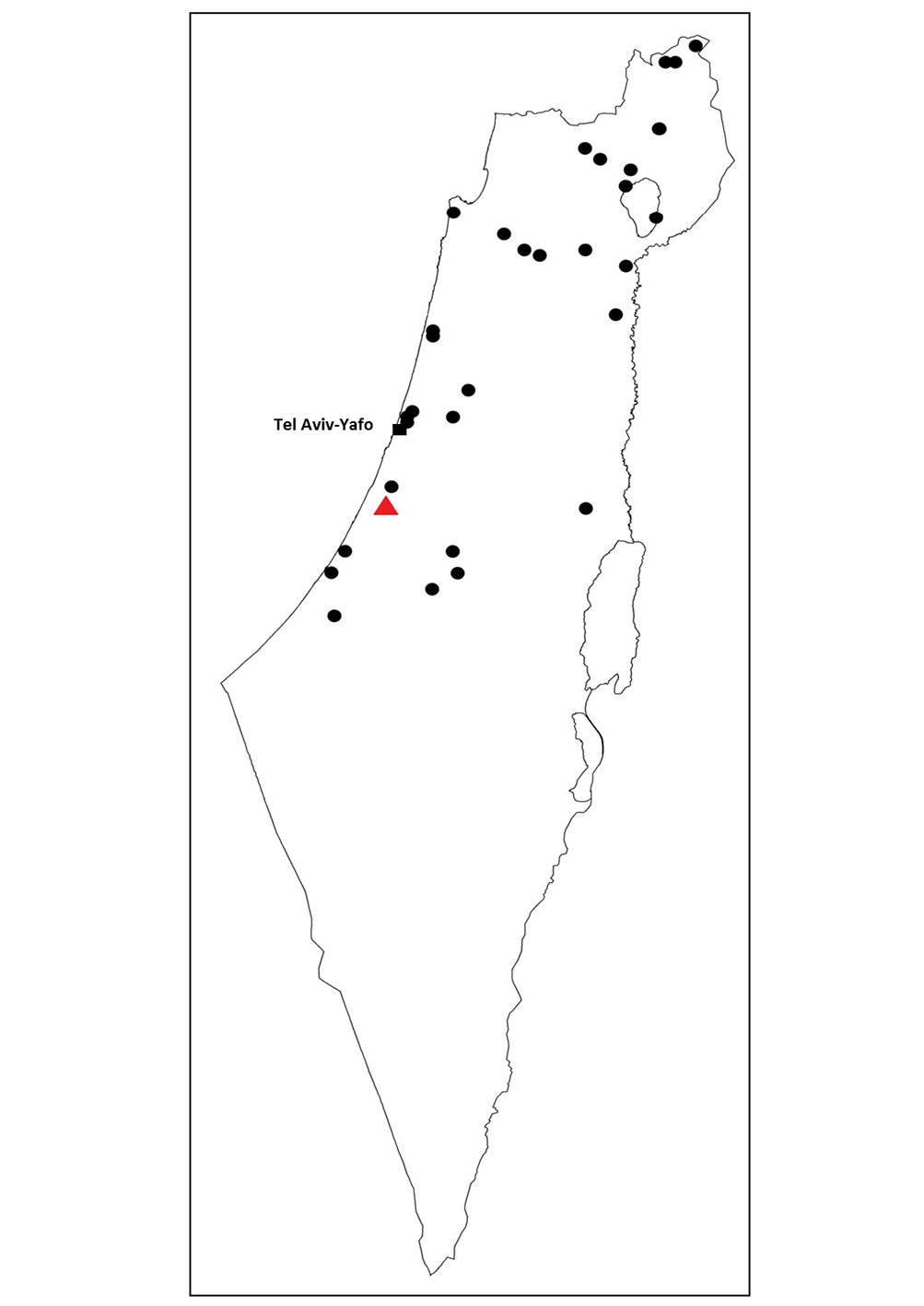
Distribution. ( Fig. 23 View FIGURE 23 ). Israel, southern Turkey.
Comments. Although the two males available from Turkey fit well the description of the new species, I refrain from including them in the paratype series due to the slight possibility that they may belong to a third species.
Etymology. The specific epithet liat is to honor my dear younger daughter, Liat , who is even more interesting and beautiful than the new species. It is a noun in apposition.







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Goniglossum liat
| Freidberg, Amnon 2016 |
Goniglossum wiedemanni
| Norrbom 1997: 340 |