
Cliona californiana
|
publication ID |
https://doi.org/10.11646/zootaxa.3823.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:0D42FA17-3B11-4DBB-9E48-D7D505F9CE29 |
|
DOI |
https://doi.org/10.5281/zenodo.6132500 |
|
persistent identifier |
https://treatment.plazi.org/id/03D0FB0A-FF8C-2E1A-09E0-FD71FB8A793F |
|
treatment provided by |
Plazi |
|
scientific name |
Cliona californiana |
| status |
|
Cliona californiana de Laubenfels, 1932
Figs. 1 View FIGURE 1 A–F; 2
Cliona celata var. californiana de Laubenfels, 1932; Pseudosuberites pseudos Dickinson, 1945
Material examined. KML 1002, KML sta. 61/69, Wharf, Bamfield E, BC, ( 48º 50.1′N, 125º 08.1′W), 5 m depth, Jun. 2, 1969, coll. W.C. Austin; KML 1003, PBS sta. 63-35, Porlier Pass, BC, ( 49º 11.0′N, 123º 41.0′W), 36 m depth, coll. D.B. Quayle (PBS); KML 1004, 10 km off Copper Bay, Haida Gwaii, BC, (approximately 53º 09′ N, 131º 36′W), no depth given, Aug. 29, 1960, coll. D.B. Quayle (PBS); KML 1005, KML sta. 16/80, Wizard Island Pass, BC, ( 48º 51.6′N, 125º 10.4′W), 15 m depth, Mar. 22, 1980, coll. W.C. Austin; KML 1006, Exeter Shoal, BC, ( 49º 39′N, 124º 39′W), 22 m depth, Jun. 28, 1962, coll. D.B. Quayle (PBS); KML 1008, Effingham Island, BC, (approximately 48º 52′N, 125º 18′W), no depth given, Jun. 6, 1961, coll. D.B. Quayle ( FRB); KML 1009, KML sta. 100/75, E. of Diana Island, BC, ( 48º 50′N, 123º 11′W), coll. W.C. Austin; KML 1010, KML sta. 50/77, Thieves Bay, BC, ( 48º 46.2′N, 123º 19.8′W), 20 m depth, Mar. 20, 1977, coll. W.C. Austin; KML 1017, KML sta. 160/75, W side of Satellite Passage, Barkley Sound, BC, ( 48o 51.1′N, 125o 10.6′W), 65 m depth, Oct. 6, 1975, coll. W.C. Austin; KML 1011, KML 85/73, Whittlestone Point, Barkley Sound, BC, ( 48º 48.5′N, 125º 11.1′W), 9–12 m, May 5, 1973, coll. W.C. Austin; KML 1189, Christie Islet, Howe Sound, BC, (49° 29.919 N, 123° 17.985W), 15 m depth, May 19, 2012, coll. & photo N. McDaniel.
Field images (not accompanied by voucher): Howe Sd., BC, (approx. 49º 28′N, 123º 20′W), photos, N. McDaniel; Flat Top Islands, BC ( 49º 9′N, 123º 41′W), photo, N. McDaniel.
Description. Macroscopic features. The basic growth form was determined for specimens from ten localities: nine were α stage, one was γ stage ( KML 1017). Colour alive bright yellow, in alcohol dark brown. Colour photos show dramatic differences between expanded and contracted sponges ( Fig. 1 View FIGURE 1 A–C).
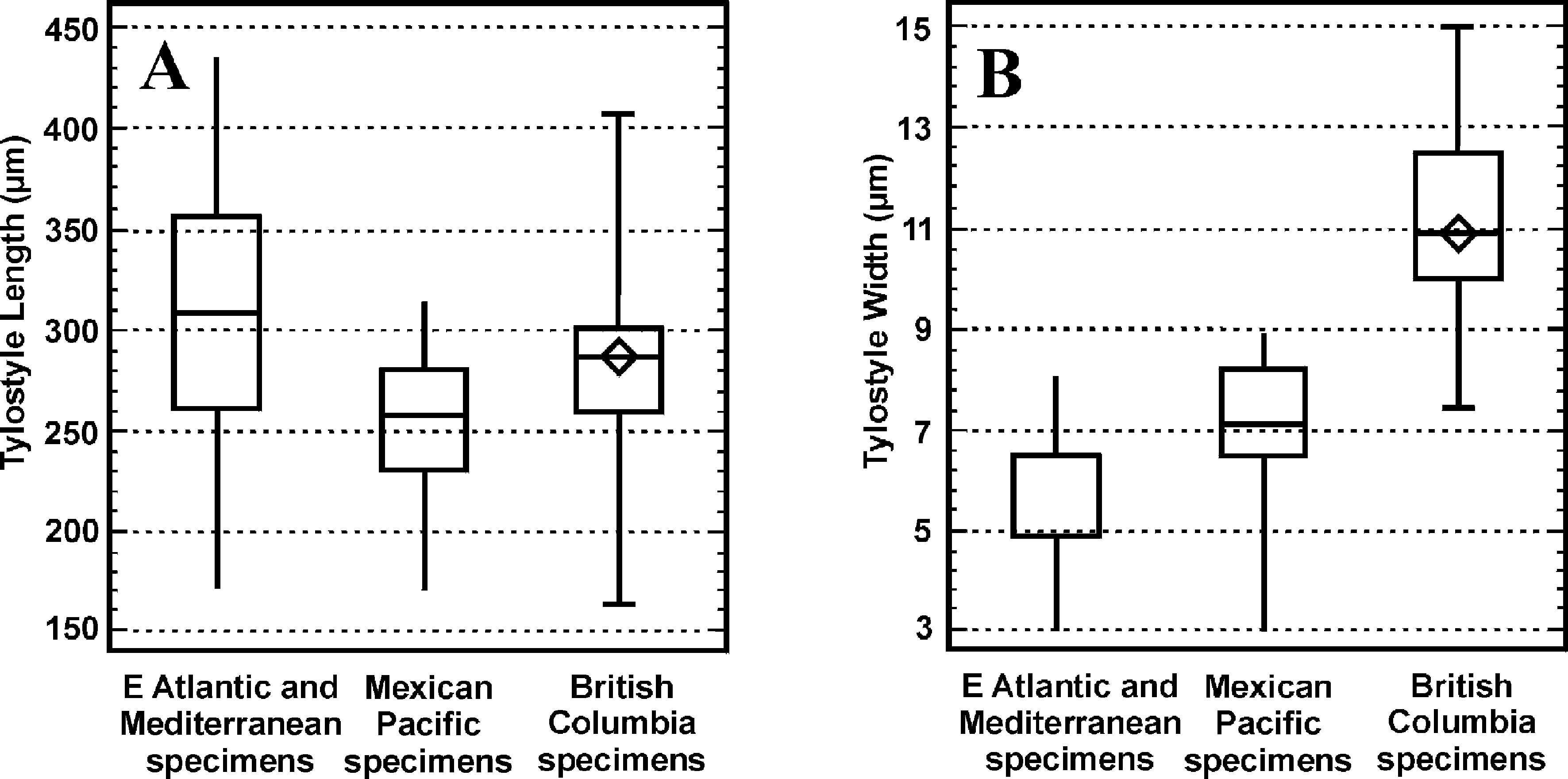
Spicules. Spicules exclusively tylostyles ( Fig. 1 View FIGURE 1 D–F), often subterminal ( Fig. 1 View FIGURE 1 F), some polytylote ( Fig. 1 View FIGURE 1 E). The range and mean tylostyle dimensions (µm) in specimens of Cliona californiana from various sites in BC are given below.
We compared tylostyle dimensions using box and whisker plots ( Fig. 2 View FIGURE 2 ) of BC specimens (our data) and of east Atlantic/ Mediterranean and Mexican Pacific specimens (data from Carballo et al. 2004).
Remarks. Carballo et al. (2004) found that Cliona celata var. californiana from the Mexican Pacific coast had tylostyles which were significantly shorter and wider than specimens of Cliona celata from the Mediterranean.
Their measure of significance was P<0.0001. F-ratio 30.04 for length and P<0.0001 F-ratio 66.98. They also noted that tylostyle heads of Mexican material had better formed tylostyles with fewer style/subtylostyle modifications than the Mediterranean specimens. They proposed that C. celata var. californiana be considered as a valid species, C. californiana , and distinct from C. celata .
When we compared our BC specimens with the box and whisker plots of Caraballo et al. 2004, we found that tylostyle lengths did not significantly vary from the Atlantic/Mediterranean or from the Mexican Pacific specimens ( Fig. 2 View FIGURE 2 A). However, tylostyles were wider in the BC specimens than in the Mexican Pacific specimens and much wider than in the Mediterranean specimens ( Fig. 2 View FIGURE 2 B). This may reflect a genetic difference sufficient to support full species status for C. californiana . However, it has also been demonstrated that in some sponges the width but not necessarily the length is directly correlated with ambient silica concentration (e.g. Stone 1970, Mercurio et al. 2000). British Columbia has one of the highest levels of silica in the world in shallow water (Austin 2012). By contrast Mediterranean waters have low silica concentrations relative to the Atlantic (e.g., Uriz et al. 2003) and the North Atlantic has low concentrations compared to the NE Pacific (e.g., Schlitzer 2000). The spicule width of Cliona celata populations could, therefore, be a reflection of differences in silica concentration.
Rosell & Uriz (2002) provided a table listing 11 populations of C. celata reported in the literature from various localities in the North Atlantic. We compared the maximum lengths and widths for each population with those for the Mexican and BC specimens. Minimums were not used as it was unclear whether or not juvenile spicules were included. Mean values were not available for most populations.
Range in maximum lengths Atlantic specimens 300–450 Μm Range in maximum lengths Mexican specimens 287–412 Μm Range in maximum lengths BC specimens 300–410 Μm Range in maximum widths Atlantic specimens 7.0–17 Μm
Range in maximum widths Mexican specimens 7.5–13.8 Μm Range in maximum widths BC specimens 12.5–15 Μm For both lengths and widths the ranges in sizes for the largest spicules of Mexican and BC specimens fit within the ranges for the Atlantic specimens except for a slightly smaller maximum size of 287 Μm compared to 300 Μm in the Atlantic populations. We also compared the ratios of length to width for the specimens. These ratios were 31 for Atlantic, 33 for Mexico, and 26 for BC. The lower ratio for the BC specimens could be an ecophenotypic response to higher levels of silica in BC (discussed above). Carballo et al. (2004) suggested that the more regular production of tylostyles with few styles or subtylostyles in the Mexican specimens compared to the Atlantic specimens supports elevation of C liona celata var. californiana to species status. However, these differences may also be ecophenotypic rather than genetic. Uriz et al. (2003) commented that the transformation of styles to tylostyles or subtylostyles may depend on silica concentration.
Conclusions. We have reservations about accepting C. celata var. californiana as a distinct species given the possibility that apparent differences in size and form of the tylostyles in Mexican specimens compared to Atlantic and Mediterranean specimens could well be ecophenotypic expressions in response to silica concentrations. Also the range in upper size limits of tylostyles for a broader selection of N. Atlantic localities ( Rosell & Uriz 2002) encompasses the range for the Mexican and BC samples. However, until more detailed data suggest otherwise, we follow Carballo et al. (2004) in considering C. californiana as a distinct species. We agree with one of the reviewers that the Cliona celata / Cliona californiana complex is a good candidate for barcoding. We extend the species range further north to British Columbia. Populations from southern Alaska are also likely to be this species. Given that we suggest barcoding and Carballo et al. (2004) have compared the Mexican specimens with those from the Atlantic, we hesitate to compound the confusion by comparing classical characters for Indo-Pacific species.
Bathymetric range. Low intertidal to 46 m depth.
Geographic distribution. British Columbia ( Canada), Washington, California ( USA), Baja California, Sonora, Sinaloa, Nayurit, Jalisco, Guerrero, Gulf of Tehuantepec (W. Mexico).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Cliona californiana
| Austin, William C., Ott, Bruce S., Reiswig, Henry M., Romagosa, Paula & G, Neil 2014 |
Pseudosuberites pseudos
| Dickinson 1945 |