
Abrawayaomys chebezi, Pardiñas, Ulyses F. J., Teta, Pablo & D’Elía, Guillermo, 2009
|
publication ID |
https://doi.org/ 10.5281/zenodo.188290 |
|
DOI |
https://doi.org/10.5281/zenodo.5616274 |
|
persistent identifier |
https://treatment.plazi.org/id/03D287C9-1815-FFEE-FF7D-BF73FC3AFC31 |
|
treatment provided by |
Plazi |
|
scientific name |
Abrawayaomys chebezi |
| status |
sp. nov. |
Abrawayaomys chebezi , new species
Figs. 1–9 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 , Tables 1–2 View TABLE 1 View TABLE 2
Holotype. MACN 20253, adult male collected by Orlando Scaglia on September 11, 1988 (original number Mi 15), preserved as skin (in regular condition), skull (partially damaged on basicranium and temporal flanks), partial postcranial skeleton, and mummified penis.
Type locality. Argentina: Province of Misiones, Department of Iguazú, conjunction Arroyo Mbocai and route 12 (- 25.680115º S, - 54.508060º W; Reig and Kirsch, 1988).
Diagnosis. A member of the sigmodontine genus Abrawayaomys smaller than A. ruschii especially in molar measurements, characterized by a unique combination of morphological traits including tail larger than head and body with a dark apical tuft; venter coloration grey cream yellowish; rostral third of nasals not conspicuously expanded laterally; nasofrontal suture V shaped, surpassing posteriorly the lacrimal level; interorbital region hourglass shaped; alisphenoid strut present; anteromedian flexus on M1 inconspicuous or absent; and upper incisors proodont.
Paratypes. None.
Other referred specimens ( Fig. 1 View FIGURE 1 ). All specimens are from the province of Misiones:
Colección Elio Massoia (CEM; Buenos Aires, Argentina) CEM s/n, craniodental remains from owl pellets; collected in Escuela Provincial 639 “Rosario Vera Peñaloza,” Lote 92, Sección II de Campo Ramón (ca. - 27.416667º S, - 55.016667º W, Department of Oberá; Massoia, 1988, 1996);
CEM 9970, anterior fragment of skull recovered from an owl pellet collected in Segunda Iglesia Cuadrangular, Barrio Parque Km. 11, Eldorado (- 26.4º S, - 54.516667º W, Department of Eldorado; Massoia et al., 1991);
CEM s/n, craniodental remains from owl pellets; collected in Montecarlo (ca. - 26.566667º S, - 54.733333º W, Department of Montecarlo; Massoia, 1993, 1996).
There is an unconfirmed published mention ( Massoia, 1993) for Puerto Caraguatay, Misiones.
Distribution ( Fig. 1 View FIGURE 1 ). Western portion of Misiones province in Argentina.
Etymology. In homage to Juan Carlos Chebez, Argentinean naturalist and friend, for his effort towards increasing our understanding and preserving the Atlantic Forest biome in the Province of Misiones, Argentina.
Morphological description based on the holotype. Medium-sized, moderately long tailed rat, with a large head ( Fig. 2 View FIGURE 2 ). The body pelage is short and close with a moderately spiny texture. There are two types of dorsal and ventral hairs of approximately the same length (1.1–1.2 mm): spiny hairs and long, thin hairs. Spiny hairs are flat, with a dorsal longitudinal groove, rigid and broadest at its midpoint (0.22 mm), especially at the dorsum. Spines are more abundant at the dorsum and rump, being slightly shorter, without dark tips, and thinner in the underparts and shorter and darker in the head; mental and inguinal areas are free of spines. Spine cuticular scales vary from imbricate with an elongate pattern at the base and middle portion of the spine and gradually transform to imbricate with a wavy crenated-flattened pattern towards the tip ( Fig. 3 View FIGURE 3 ). General appearance is strongly agouti and hispid, darker at the midline and without a clear separation of the dorsal and ventral color. Dorsal and lateral spiny hairs are slightly transparent or whitish at the base and throughout almost their entire length, becoming brownish toward the tip. Thin hairs are brownish at basal portion, with buffy subterminal portions and apical portion. Intermixed with spines and thin hairs there is an unusual type of hair, for which we could not find a name in the literature, composed by a bouquet of thin, dark grey hairs arising from a unique root ( Fig. 4 View FIGURE 4 a). Chin and the interior sides of the fore- and hind feet are covered by graybased buffy hairs. On the underparts, spines and fur are silvery-transparent at the base, becoming buffy toward the middle and apical portions. Ears are small and rounded, internally covered by short golden hairs and externally by brown hairs. The dorsal part of the muzzle is slightly lighter than the rest of the head. Vibrissae are long and thin; mystacial, supercilliary, and genal vibrissae reach the mid-portion of the pinnae; all vibrissae are buffy-brown colored. There is also a genal vibrissa 2 (sensu Pacheco, 2003:32). The tail is uniformly brown with conspicuous scales and a short (ca. 8 mm) tuft of dark brown hairs in the tip ( Fig. 4 View FIGURE 4 b); tail hairs are rigid and dark and are arranged in sets of 5 per scale; each set are composed by 3 central hairs subequal in length flanked side by side by 1 shorter hair; tail hairs are about 1 scale-row long at the base, 1 ½ scale-rows long at the mid-portion, and 3 scale-rows long near the tail end ( Fig. 4 View FIGURE 4 c). The manus are whitish, with long digits ( Fig. 4 View FIGURE 4 d); the three interdigital pads are well defined and rounded; the hypothenar and thenar pads are large and narrow; manual DIII and DIV are nearly equal in length, and DII and DV are subequal with DII slightly longer; claws are short (1.7 mm), curved and narrow; the surrounding tufts of hairs reach or surpass claw tips. The pes are withish, long and narrow ( Fig. 4 View FIGURE 4 e), with five large digits distally appearing bicolored due to the abundance and the whitish color of the ungual tufts ( Fig. 4 View FIGURE 4 f); DII and DIV are subequal, DIII is slightly longer than the previous ones; DV reaches the middle of the second phalanx of DIV; DI (hallux) is short, level with the middle portion of the second interdigital pad and has a claw; four ovale interdigital pads are present; hypothenar and thenar are large, rounded and narrow; the distal margin of the former plantar pad is leveled with the proximal margin of the second; the plantar surface is smooth and naked, including the heel; pedal claws are short and ungual tufts surpass the claws in length ( Fig. 4 View FIGURE 4 f).
The skull is robust, with a short and blunt rostrum, wide interorbital region and a nearly square braincase ( Fig. 5 View FIGURE 5 ). The nasals are narrow, parallel-sided in the proximal half, with a marked abrupt expansion in the anterior one third, and shorter than the premaxillae, leaving visible the anterior part of the later and the incisors in a dorsal view; posterior end of nasals are extended beyond the premaxillary-maxillary suture ( Fig. 6 View FIGURE 6 a). The narines are wide and the turbinals strongly developed ( Fig. 6 View FIGURE 6 b). The nasofrontal suture is slightly acuminate and almost without denticulations, positioned surpassing posteriorly the line defined by the lacrimal bones ( Fig. 6 View FIGURE 6 c). The lacrimals are large, in contact with the maxillaries and the frontals. The anterior part of the interfrontal region has a moderate central depression also extended to the internasal one. The interorbital region can be typified as “hourglass-shaped,” although is relatively broad producing a partial dilution of the constriction point; the supraorbital margins are rounded ( Fig. 6 View FIGURE 6 c). The coronal suture is wide and U-shaped ( Fig. 6 View FIGURE 6 c). Lateral parietal crests are slightly marked, but lambdoid suture is concealed by a definite lambdoidal crest where the squamosal meets the occipital bone ( Fig. 5 View FIGURE 5 ). The parietal and the interparietal sutures are meandrous; the interparietal is wide and broad, rhomboidal in form. The occipital region is squared in lateral view, producing a nearly vertical position of the foramen magnum. The zygomatic plate is high, parallel-sided and conspicuously broad from top to bottom, with a dorsal free margin and a nearly vertical anterior border; its posterior margin is situated anterior to the alveolus of M1 ( Fig. 6 View FIGURE 6 d). Zygomatic notches are wide and nasolacrimal capsules are clearly visible in dorsal view. The zygomatic arches are well flared and partially dorso-ventrally expanded in their midpoints; the jugal is large and the maxillary and squamosal processes of the zygoma are not in contact ( Fig. 6 View FIGURE 6 e). The squamosal process of the zygomatic arch supports an anterior-posteriorly expanded glenoid cavity. Anteriorly, the periotic is attached to the skull by a well developed tegmen tympani, which overlaps the posterior suspensory process of the squamosal (sensu Voss, 1993:18). The hamular process of the squamosal is broad, distally spatulated and directly applied to the mastoid tubercle; the postglenoid foramen and the subsquamosal fenestra are well developed and sub equal in size ( Fig. 6 View FIGURE 6 g). The parietal has a moderate lateral expansion subrectangular in form ( Fig. 6 View FIGURE 6 g). The osteological structures associated to the carotid circulatory pattern include a well developed squamosal-alisphenoid groove that anteriorly leads to a small sphenofrontal foramen; a shallow trough for masticatory-buccinator nerve connects to a small foramen ovale accesorius ( Fig. 7 View FIGURE 7 c); these traits, along with the presence of a groove moderately expressed on the posterior external edge of the pterygoid plate ( Fig. 6 View FIGURE 6 f), and the presence of the stapedial foramen on auditory bullae, suggest that Abrawayaomys possess the presumably primitive muroid condition (type 1 of Voss, 1988) for the cephalic blood supply (see also Pereira et al., 2008). A well developed alisphenoid strut crossing the foramen ovale accesorius is present ( Fig. 7 View FIGURE 7 ). The incisive foramina are short, not reaching the alveolus of the M1, and narrow, with straight and parallel lateral margins. The palate is, by definition, wide and long (sensu Hershkovitz, 1962), although the anterior margin of the mesopterygoid fossa is only slightly posteriorly placed with respect the plane defined by the M3 posterior faces ( Fig. 6 View FIGURE 6 f). Three paired small posterior palatine foramina between the upper molars and minute unpaired posterolateral palatal pits side by side the anterior part of the mesopterygoid fossa ( Fig. 6 View FIGURE 6 f) are present. The mesopterygoid fossa is large and wide, with a rounded U-shaped anterior border and its roof completely ossified ( Fig. 6 View FIGURE 6 f). The parapterygoid fossae are narrow and well ossified. The periotic capsule of the petrosal is large and moderately inflated, without fenestrae. The ectotympanic bullae are globular capsules of moderate size, being the periotic partially exposed; the auditory meatus are rounded. The manubrium of the malleus is large and distally expanded; a well developed orbicular apophysis is present (“parallel” type, according to Carleton [1980]).
The mandible is relatively short, high, and robust ( Fig. 5 View FIGURE 5 ). The angular process is short with respect to the condyloid process ( Fig. 6 View FIGURE 6 h). The coronoid process is triangular in shape, short, and posterodorsally oriented. The sigmoid notch is narrow and subelliptical. The lunate notch is shallow ( Fig. 6 View FIGURE 6 h). The capsular projection of the lower incisor lies under the angular notch and is marked, forming a distinct knob in dorsal view ( Fig. 6 View FIGURE 6 h). The anteriormost point of the diastema is below the alveolar plane ( Fig. 6 View FIGURE 6 i). The mental foramen is visible from lateral view, just in front of the conjunction of the upper and lower ridges of the masseteric crests ( Fig. 6 View FIGURE 6 i); both the upper and lower ridges of the masseteric crest are well marked. The ventromedial process of the mandibular ramus is only slightly expressed. The retromolar fossa is long and wide, moderately foraminated ( Fig. 6 View FIGURE 6 j). Internally, a well developed and curved duct for the mandibular nerve is visible through the translucent bone and leads to a large mandibular foramen ( Fig. 6 View FIGURE 6 k).
The upper incisors are proodont ( Fig. 6 View FIGURE 6 d), markedly deeper but transversally compressed ( Fig. 6 View FIGURE 6 l) and have deep, orange frontal enamel; the dentine fissure is straight and large (sensu Steppan, 1995:23). The general morphology of the upper incisors suggests their allocation under the seizer-digger type (sensu Hershkovitz, 1962:102); supporting this assignment can be mentioned the following features: cross section above the cutting edge (anteriorly convergent), the acute angle with respect to the sagital plane of the skull, and their narrowing towards the tips ( Fig. 6 View FIGURE 6 l). The lower incisors are noticeably deep but transversally thin and the frontal enamel paler than upper one ( Fig. 6 View FIGURE 6 i).
The upper molars are brachyodont and the coronal surface slightly crested being the lingual cusps something taller than the labial ones. The main cusps arrangement is intermediate (sensu Steppan, 1995:24; Fig. 8 View FIGURE 8 a). In the M1 the procingulum is anterior-posteriorly compressed and labially displaced. There is no evidence of anteromedian flexus on the enamel anterior wall but an inconspicuous dentine indentation can be ascribed to this structure. The procingulum is lingually projected by a presumptive protostyle and a cingulum that externally reaches the protocone enclosing the protoflexus. The general morphological configuration and orientation of the procingulum of the M1 are achieved by a reduced anterolingual conule and a presumptive parastyle fused and projecting the anterolobial one. The areas of proto- and hypocone are subequal and bulbous; the same is true for the areas of para- and metacone. The hypocone is slightly labially recessed with respect to the protocone; the hypoflexus is wide, scarcely penetrates the molar midline, and has a corrugated base. The lingual margin of the M1 between proto- and hypocone is ridged by a cingulum that closes the hypoflexus transforming it into a basin. Although wear erases occlusal structures in the intermediate area between para- and metacone, an indication of a narrow but long mesoloph fused with a paralophule is distinguishable. At least in this wear stage, the posteroloph is fused to the metacone. The M2 general configuration is very similar to the comparable portion of the M1. The procingulum is well developed, labially and lingually closed by sharp cingula. The hypocone is also recessed with respect to the protocone; a presumptive enterostyle is present. The mesoloph-paralophule development is like those in M1. A posteroloph directly backwards is independent of the metacone. The M3 is notably reduced, cylindrical in outline, and seems to have only one root. The occlusal morphology suggests the persistence of solely the proto- and paracone areas, being the posterior portion of this tooth deeply compressed.
The lower molars, also brachyodont, have the coronal surface crested like the upper molars; the lingual cusps are taller than labial ones and arranged in a clear alternate pattern (sensu Steppan, 1995:24; Fig. 8 View FIGURE 8 b). The enamel is thick but there is a good amount of exposed dentine. In the m1 the procingulum is clearly transversally compressed and remains as a narrow structure on the labial sector of the tooth without evidence of an anteromedian flexid. The proto- and hypoconid look like enamel cusps without wear; meta- and entoconid are bulbous and subequals in size. The hypoflexid is wide, short, and slightly labially closed by a weak cingulum. On the posterior face of the metaconid a short mesolophid is present. The posterolophid is well developed. One of the most peculiar characteristic in the m1 morphology is the compression displayed by the lingual cusps and their projection like peninsulas on the lingual margin. The m2, slightly shorter than m1, is rectangular in outline and has the basic pattern of the m1. The posterolophid is short and directed backwards. The m3 is comparatively reduced but not as the upper one, with a greatly reduced entoconid and a posterior enamel lake produced by the obliteration of the posteroflexid.
Axial skeletal counts include 12 ribs, 15 thoracicolumbar (dorsal) vertebrae, 3 sacral vertebrae, and 36 caudal vertebrae. The neural spine on the second thoracic vertebra is much longer than the spines on the adjacent vertebrae (a widespread condition for sigmodontines according to Voss, 1988). Hemal arches are present between tail vertebrae second and third, and between third and fourth; regrettably, hemal arch presence between the first and second caudal vertebrae in impossible to check due the incomplete nature of the skeleton under study. The posterior edges of the hemal arches are extended into a distinct spinous process (sensu Steppan, 1995:49), less developed between third and fourth. Hemal processes starts on fourth vertebrae, becoming most pronounced between fifth and sixth, and then diminishing along the next ten vertebrae. The tuberculum of the first rib articulates with the transverse process of both the seventh cervical and the first thoracic vertebrae. The sternum consists of an anterior manubrium, four sternebrae, and a posterior xiphoid process. The humerus lacks an entepicondylar foramen; the supratrochlear fossa is perforated. The innominate has a well developed femoral tubercle positioned cranially to the acetabulum; the acetabulum is small and rounded; the inferior and superior glutear fossa of the ilium are divided by a rounded lateral ridge. The femur has a small head and a long and narrow patellar fossa; the condyles are slightly rounded in contour.
The gland penis is short, stocky and covered by spines throughout the entire body. The mid-shaft diameter is slightly less than one-half of the overall length. Dorsal and ventral grooves are deep and extend beyond half the length of the body. The crater rim is apparently uniform; but unfortunately the apical portion of the penis, including the bacular mounds, is damaged precluding the possibility of an accurate description. In appearance, lateral bacular mounds are not visible in external view; the medial bacular mound is slightly larger than the lateral ones. The proximal baculum is short, stout, and triangular in appearance, with a laterally flared base. The base has a distinct and deep median notch. The shaft is short and wide. The cartilaginous distal baculum is small, with two laterals and one medial digit.
Measurements. External and craniodental measurements of the holotype of A. chebezi , new species, are provided in Table 1 View TABLE 1 along with those from other specimens of Abrawayaomys from Brazil.
Habitat. Recording localities of Abrawayaomys in Argentina are placed in the Paraná-Paraíba interior forest ( Olson et al., 2001) of the Misiones Province. Specimen MACN 20253 was captured using a Sherman trap in the southern margin of the arroyo Mbocaí, in a secondary forest belonging to “Rosewood and Assai Palm Forest” unit (sensu Martínez-Crovetto, 1963), where the predominant tree species are Matayba elaeagnoides , Begonia descoleana , and Podostemun comatum. The remainder Argentinean specimens were recovered from Tyto alba owl pellets and came from areas that are highly disturbed by human activities, where only small patches of the “Laurel and Guatambu forest” survives. It is interesting to note that Campo Ramón is placed near the northern border of the natural grassland ecoregion of “Campos.” Although small gallery forest fragments persist in this ecoregion, the extensive conversion to agriculture landscapes is noticeable ( Giraudo et al., 2002). It is also interesting to remark that collection localities for Abrawayaomys in Misiones are located on the western flank of the province that drains towards the Paraná River. No record is known from the eastern flank, which drains towards the Uruguay River. However, is important to remake that trapping effort on the Uruguay basin is clearly lesser than that on the Paraná basin.
Comparisons. Although the small amount of available specimens precludes a thoroughly assessment of intra and interpopulation variation, some comments can be advanced from individual measurements ( Table 1 View TABLE 1 ). The holotype (MACN 20253, Misiones, Argentina) of Abrawayaomys chebezi , new species, and an additional specimen (CEM 9970, Misiones, Argentina) reported by Massoia et al. (1991) seem to be slightly smaller in most of their cranial dimensions than A. ruschii holotype (MN 23075, Espírito Santo, Brazil) and those specimens from Brazil that represent the northern part of the range of the genus. This fact is particularly noticeable in the length of the upper molar tooth-row, incisive foramina, and mandible. By contrast, specimen MACN 20253 has the rostral length clearly larger than those of Brazilian specimens despite similar condylobasal lengths among all individuals. Trenchant differences in molar individual measurements arise when MACN 20253 and MN 23075 (data after Cunha and Cruz, 1979:5; in mm) are confronted, respectively: M1: 1.82 vs. 2.62; M2: 1.32 vs. 1.87; M3: 0.63 vs. 0.93; m1: 1.82 vs. 2.29; m2: 1.45 vs. 1.91; m3: 0.89 vs. 1.45.
The contrast in the tail length and other traits of this organ across the available specimens deserves special mention. The tail length of the holotype of A. ruschii (MN 23075) was reported by Cunha and Cruz (1979:2) as 85 mm, being clearly shorter than head and body length (116 mm; erroneously indicated in Cunha and Cruz [1979:2] as “cabeça e corpo [head and body]: 201” that must be interpreted as the total length); other Abrawayaomys specimens from Argentina and from the Brazilian states of Santa Catarina and Minas Gerais have tails larger than head and body lengths (cf. Table 1 View TABLE 1 ). Judging by the tail tip figured by Cunha and Cruz (1979:fig. 3), can be presumed that the holotype exhibit an incomplete tail. However, these authors clearly stated that “A cauda é mais curta que a cabeça e o corpo, com pouco pelo [sic]... e um pincel na extremidade” ( Cunha and Cruz, 1979:3), freely translated “the tail is shorter than head and body, less haired... and [with] an a After Massoia et al. (1991:40);
b After Pereira et al. (2008:table 1);
c Still unnumbered specimen; data from Maria Olímpia Garcia Lopes (pers. comm.); d Data from Cherem et al. (2005:110), UFSC = Universidade Federal de Santa Catarina, Santa Catarina, Brazil; e Following toothwear classes proposed by Voss (1991);
f From dry skin; original measurements on specimen label [tail = 70, hind foot = 34, and ear = 35.6] are clearly wrong; g Partial, according to Pereira et al. (2008:table 1);
h Cunha & Cruz (1979:5) reported 5.42.
apical tuft.” João Oliveira, curator from Museu Nacional, kindly examined for us the tail of the holotype; he obtained a value of 95 mm from the dry skin and could not find any indication of tail cutting and discarded the hypothesis of an individual with a tail cut during life and later cicatricized. From all these data we can state that typical A. ruschii has a tail shorter than head and body length (ratio tail length/head and body length = 0.73). All other measured specimens have tails larger than head and body ( Table 1 View TABLE 1 ), accounting in A. chebezi , new species, a ratio of 1.10. Regrettably, the length of the tail in the MN 67557 (Rio de Janeiro, Brazil) is not available.
Belly coloration Grey White White Grey White
TL/HBL 1.1 Not available 0.8 1.1 1.1 Other contrasting tail trait is the apical tuft. While in A. chebezi , new species, the tuft is moderately short and composed by brown dark stiff hairs, in the A. ruschii holotype the tuft is almost non-existent, whereas in Minas Gerais specimens (e.g., UFMG 2492, USNM 552416, and a recently collected and still unpublished specimen [J. Oliveira, pers. comm.]) the tuft is well haired with large white or white-cream in coloration elements. More indeed, the ending one inch of the tail of the UFMG 2492 is entirely white all-around.
Specimens MACN 20253 and MN 67557 (see Pereira et al., 2008:fig. 2) have an acute naso-frontal suture surpassing posteriorly the line defined by the lacrimal bones. By contrast, in specimens from further northern localities (UFMG 2492, USNM 522416) the nasals are abruptly truncated at or near the lacrimal plane having a straight naso-frontal suture. These latter specimens also shown the anterior tips of the nasals more flared and upturned ( Fig. 9 View FIGURE 9 ).
More intriguing are the differences reported in molar morphology, particularly in the M1 procingulum. According to Cunha and Cruz (1979:3) and to our direct inspection, the holotype of A. ruschii (MN 23075) has an anteromedian flexus dividing the procingulum in subequals anterolabial and anterolingual conules. Pacheco (2003:70) also noted the presence of an anteromedian flexus in the specimen USNM 522416 (Minas Gerais, Brazil), although he indicated an anterolingual conule smaller than the anterolabial one. However, in an additional specimen from Minas Gerais (UFMG 2492) there is no trace of anteromedian flexus. Either MN 67557 (Rio de Janeiro, Brazil), and MACN 20253 (Misiones, Argentina) lack the anteromedian flexus or it has reduced to a minimum expression, respectively. Clearly, this trait is partially age-dependent but when the anteromedian flexus is well expressed in young individuals, at least an indication of it, usually as a shallow notch in the anterior face of the M1, remains in adults.
Finally, some degree of variation was observed with respect to the presence of the alisphenoid strut. Although reported by Pereira et al. (2008:35) as absent in the specimen MN 67557 (Rio de Janeiro, Brazil), our suspicion of their presence, based on a detailed inspection of the figure provided by these authors, was confirmed by João Oliveira through a direct study. In fact, only the specimen UFMG 2492 (Minas Gerais, Brazil) lacks the alisphenoid strut that is present in all the remainder specimens examined ( Fig. 7 View FIGURE 7 ).
Perhaps the more marked differences arise when the specimen USNM 522416 (Minas Gerais, Brazil) and MACN 20253 (holotype of A. chebezi , new species, Misiones, Argentina) are directly compared to. The former has partially convergent interorbital region (hourglass in the latter), shorter rostrum (longer), well on inflated nasolacrimal capsules (less conspicuous), more rounded braincase (more squared), rhomboidal and smaller interparietal (oblong and larger), longer and more robust molars (shorter and slender), broader incisive foramina (narrower), sphenopalatine vacuities present as fissures (absent, roof of mesopterygoid fossa completely ossified), large posterior openings of the alisphenoid canal on parapterygoid plates (small), and slightly opisthodont upper incisors (proodont). In addition, specimen USNM 522416 has the tail tuft white while that of MACN 20253 is dark brown.
Differences previously commented are summarized in Table 2 View TABLE 2 . We are confident that those specimens from the southernmost part of the range of the genus (Misiones, Argentina) reflect a new species here described as A. chebezi . Regarding the remaining specimens from Brazil, the biological significance of these differences, if there is any significance apart from individual variability, is hard to evaluate without voucher specimens at hand. Specimens from Argentina and Rio de Janeiro appear to be more similar (maybe conspecific?) and, morphologically depart less from A. ruschii holotype than from those from Minas Gerais. Taking in mind the amplitude of the morphological variation observed across the entire Abrawayaomys sample, a coordinate treatment for specimens from Minas Gerais (e.g., UFMG 2492, USNM 552416) would be their placement on a new specific entity. The adequacy of this action should be tested and implemented, if needed, by a future reviewer, based on larger series.
The existence of more than one species, two or maybe three, in this poorly known cricetid cannot be considered as an unforeseen event on sigmodontine history. We should remember that sympatric and ecologically similar sigmodontines, which for decades were considered monotypic, recently were shown to have a moderate specific diversity (e.g., Juliomys , Rhagomys ; Luna and Patterson, 2003; Costa et al., 2007; Pardiñas et al., 2008).
No much can be said about the processes behind the diversification of Abrawayaomys ; however, its species seem to be geographically delimited by the main montane (Serras) systems of south-eastern Brazil ( Fig. 1 View FIGURE 1 ). Given this, cladogenetic events within Abrawayaomys may have been occurred in isolation on montane forest fragments prompted by climatic changes (e.g., Grazziotin et al., 2006; Cabanne et al., 2007; Carnaval and Moritz, 2008). This hypothesis, loosely framed in the Montane Isolate Hypothesis ( Moreau, 1966; Roy, 1997; see also Lara et al., 2005), should be tested with the analysis of additional samples once the specific status of populations from Minas Gerais has been clarified.
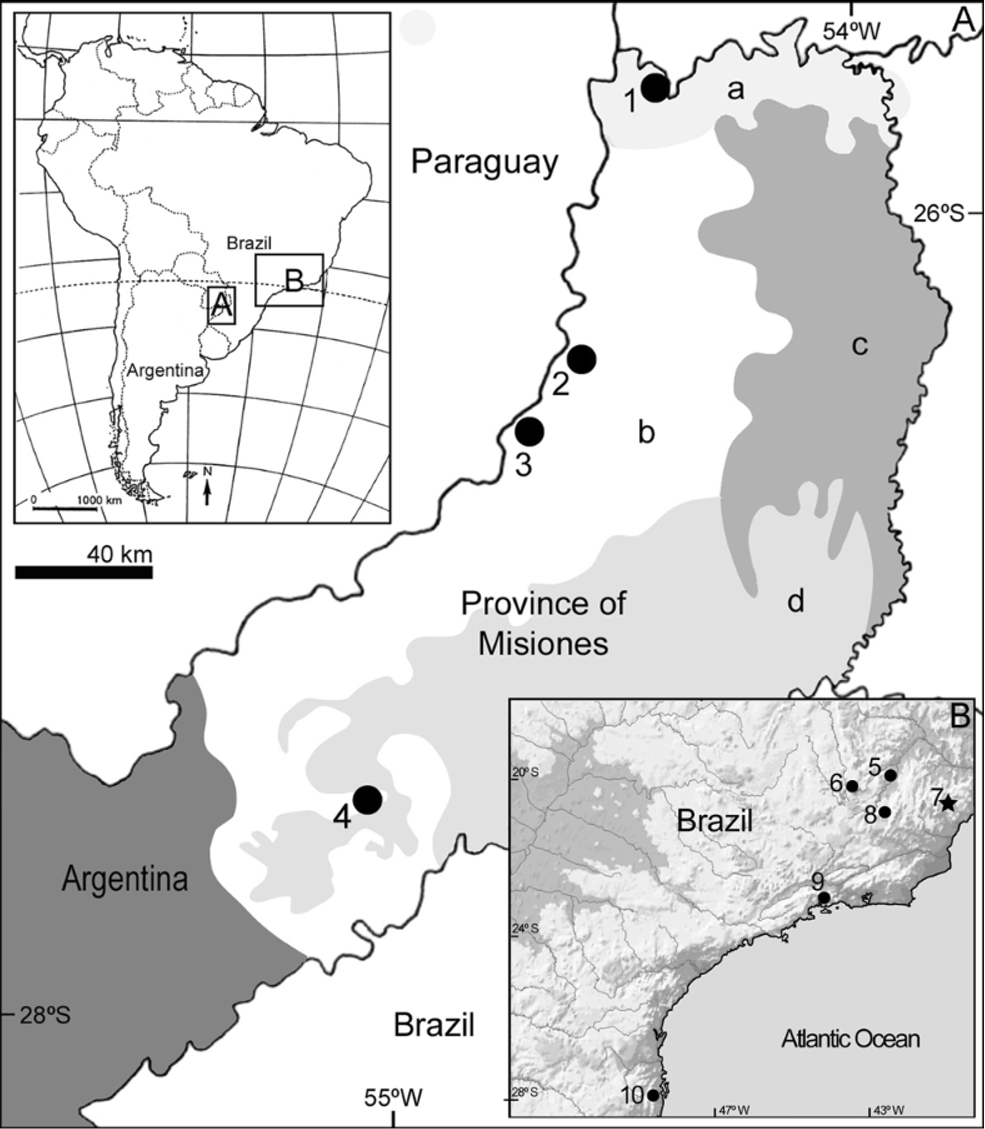
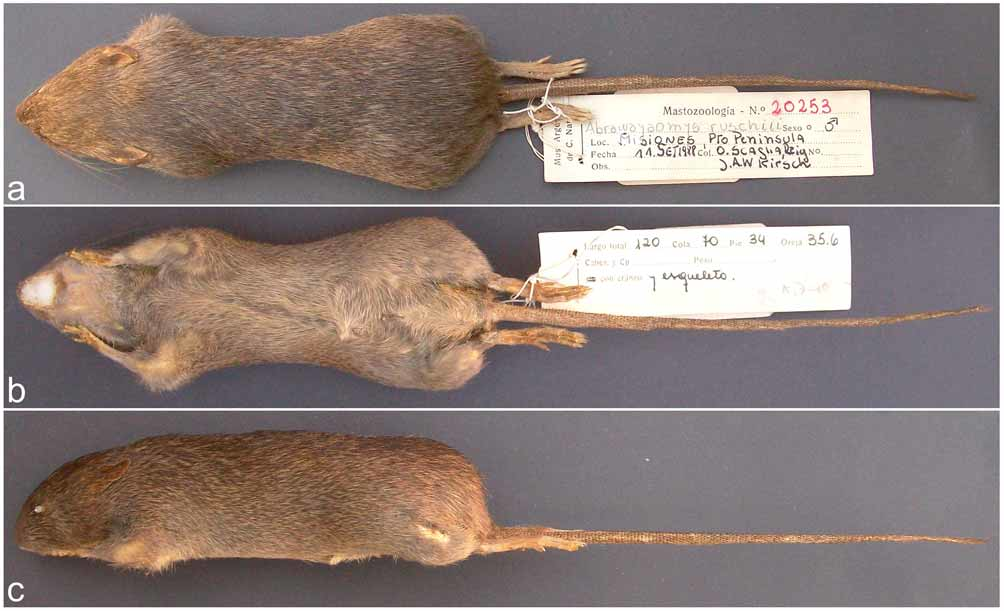


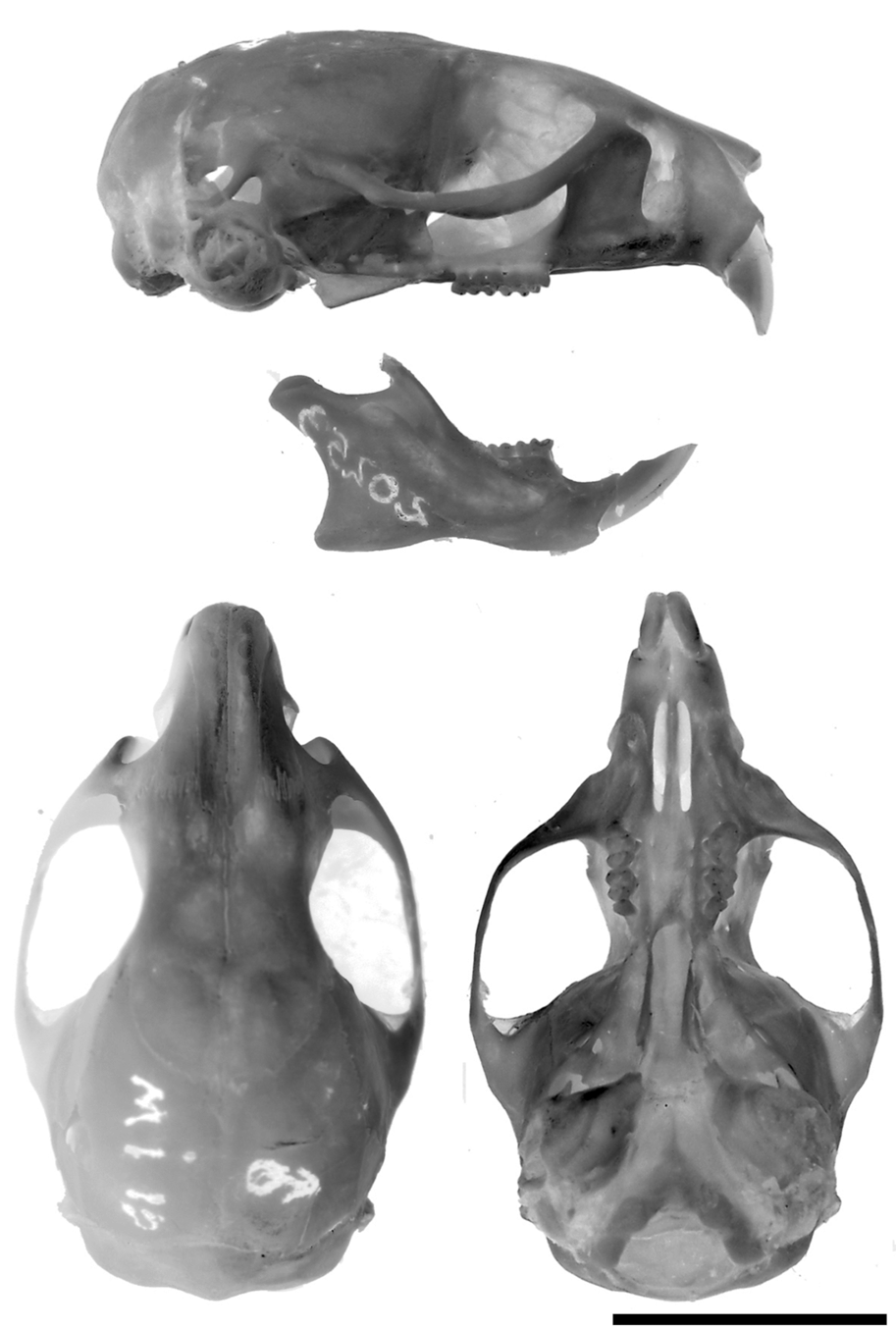
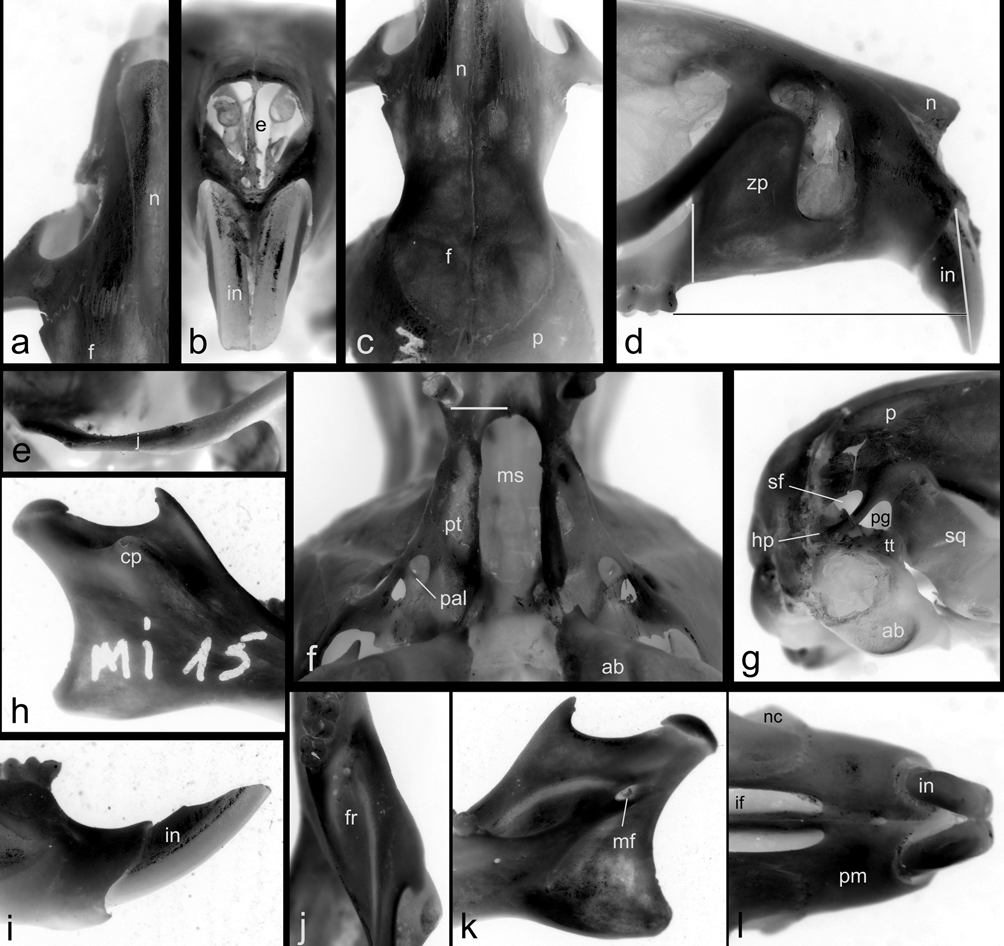
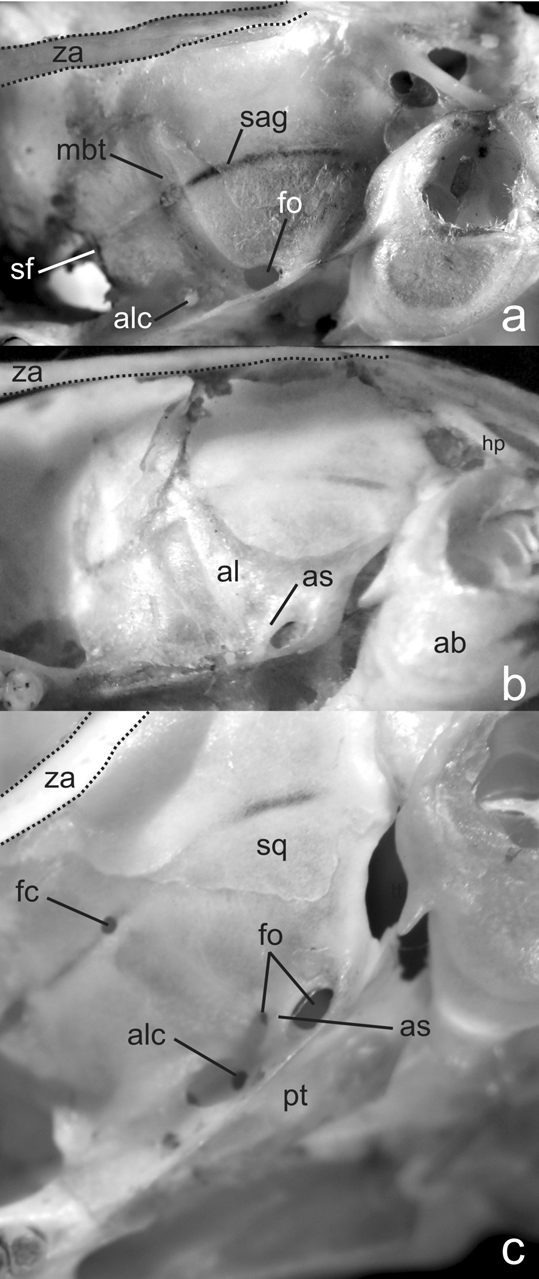
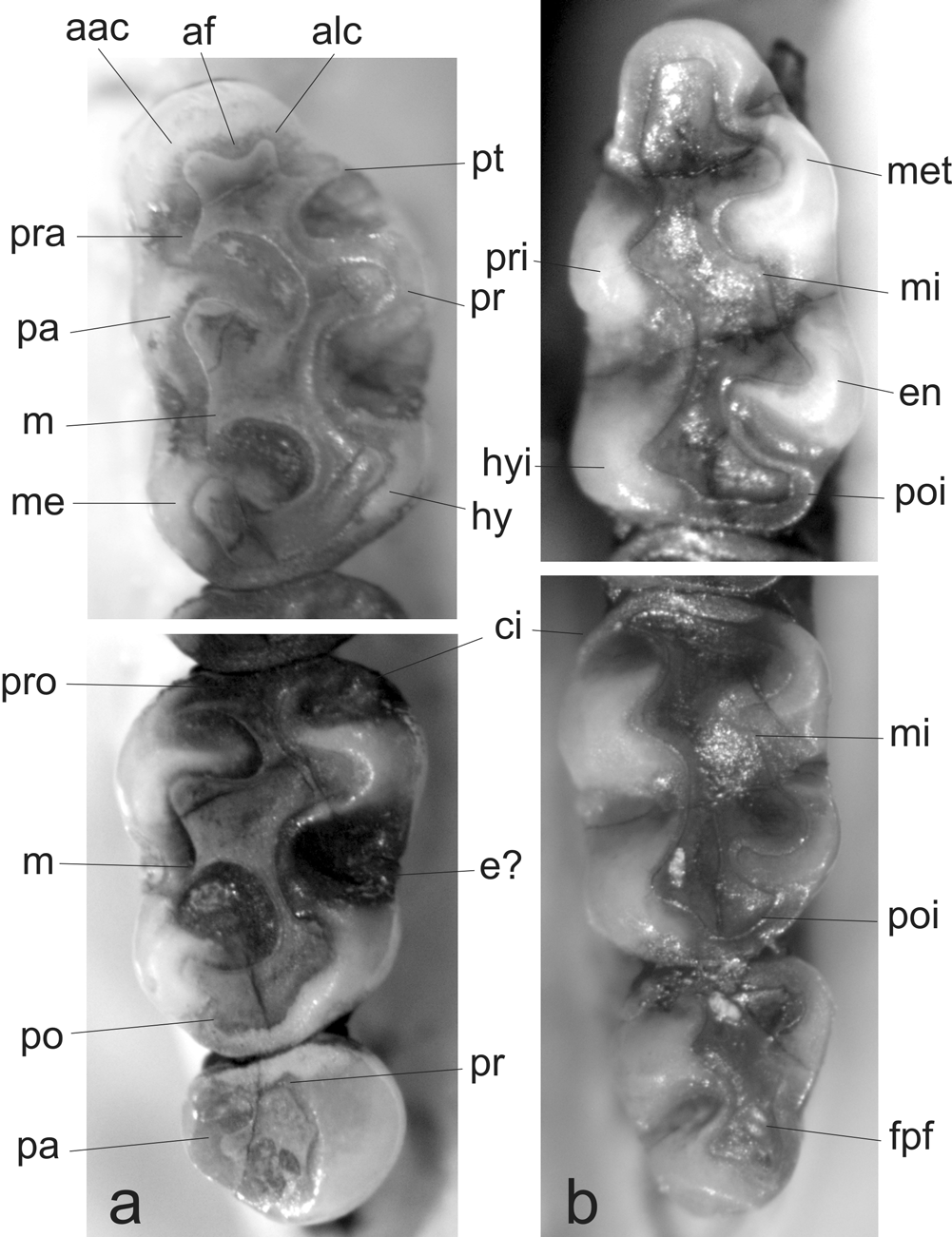

TABLE 1. External and craniodental measurements for specimens of Abrawayaomys.
| CEM 9970a | MACN 20253 holotype | MN 23075b holotype | MN 67557b | MN c | UFMG 2492b | UFSC 3427d | USNM 552416 | |
|---|---|---|---|---|---|---|---|---|
| Sex | - | male | female | male | male | male | male | male |
| Age class e | - | 4 | 3 | 3 | ? | 4 | ? | 3 |
| Head and body length [HBL] | - | 120 | 116 | 135 | 122 | 128 | 116 | 140 |
| Tail length [TL] | - | 133f | 85 | 116g | 144 | 146 | 124 | 150 |
| Hind foot length without claw | - | 27.5 | - | - | - | - | 30 | - |
| Hind foot length with claw | - | 29f | 29 | 32 | 30 | 31 | 32 | 30 |
| Ear length Weight (in grams) | - - | 17f - | 20 46 | 16 55 | 20 59 | 20 63 | 18 42 | - - |
| TL/HBL | - | 1.10 | 0.73 | - | 1.18 | 1.14 | 1.06 | 1.07 |
| Condylobasal Occipital condyle width | - - | 27.67 6.53 | 27.46 8.01 | 28.64 8.00 | - - | 27.64 7.07 | - - | - - |
| Diastema | - | 7.68 | 7.57 | 8.43 | - | 7.97 | - | - |
| Palatal bridge Incisive foramina length | - 4.40 | 5.25 4.65 | 5.46 5.33 | 5.21 5.56 | - - | 5.87 4.83 | - - | - - |
| Incisive foramina width | 1.20 | 1.44 | 1.72 | 1.63 | - | 1.68 | - | - |
| Upper molar row length | 3.30 | 3.96 | 4.51h | 4.25 | - | 4.62 | - | - |
| M1 breadth | - | 1.16 | 1.32 | 1.29 | - | 1.39 | - | - |
| Breadth between M1 Bullar length less tube | - - | 5.32 4.84 | 5.67 5.48 | 5.33 5.34 | - - | 5.82 4.86 | - - | - - |
| Cranial height | 11.00 | 9.68 | 9.94 | 10.34 | - | 10.22 | - | - |
| Rostral length Rostral width | - 5.60 | 10.94 5.62 | 8.26 5.04 | 8.41 5.34 | - - | 9.15 5.44 | - - | - - |
| Least interorbital breadth | 5.90 | 6.02 | 6.29 | 6.21 | - | 6.23 | - | - |
| Internal orbital length Zygomatic breadth | - 16.00 | 9.81 16.99 | 10.10 17.20 | 11.06 17.40 | - - | 10.70 16.00 | - - | - - |
| Braincase width | - | 13.67 | 13.18 | 13.57 | - | 13.08 | - | - |
| Zygomatic plate width Mandible height | - - | 3.56 9.03 | 4.34 9.18 | 4.31 9.23 | - - | 3.70 9.45 | - - | - - |
| Mandible length | - | 16.56 | 17.30 | 17.70 | - | 17.41 | - | - |
TABLE 2. Comparisons among Abrawayaomys specimens using selected morphological traits. MACN MN MN UFMG USNM 20253 67557 23075 2492 552416
| Tail apical tuft | Inconspicuous, dark | Not available | Very short | C o n s p ic u o u s, white | Conspicuous, white |
|---|---|---|---|---|---|
| Na sals posterior extension | Surpassing lacrimal level | S u r p a s si n g lacrimal level | Lacrimal level | Lacrimal level | Lacrimal level |
| Interorbital region morphology | Amphoral | Amphoral | Convergent | Convergent | Convergent |
| Alisphenoid strut | Present | Present | Present | Absent | Present |
| Foramen ovale on pterigoyd plate | Small | Small | ? | Large | Large |
| U p p e r in c is o r s orientation | Proodont (ca. 95º) | Proodont (ca. 95º) | S l ig h t l y opisthodont (ca. 88º) | Opisthodont (ca. 86º) | Orthodont (ca. 90º) |
| A n te r o m e d ia n flexus on M1 | Inconspicuous | Absent | Conspicuous | Absent | Conspicuous |
| MACN |
Museo Argentino de Ciencias Naturales Bernardino Rivadavia |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.