
Orussus sayii, Westwood, 1830
|
publication ID |
https://doi.org/10.1046/j.1096-3642.2003.00080.x |
|
publication LSID |
lsid:zoobank.org:pub:F3A71D74-A170-4F53-B55E-84C23D063C53 |
|
persistent identifier |
https://treatment.plazi.org/id/03D3878E-5101-7262-FB6D-BD31E34EFE54 |
|
treatment provided by |
Carolina |
|
scientific name |
Orussus sayii |
| status |
|
Distribution. Orussus is the largest and most widespread genus of Orussidae . Most known species are found in the northern hemisphere (Palearctic: 8; Nearctic: 5; Oriental: 4), including the most widely distributed species of all, O. abietinus , which ranges throughout most of the Palearctic from Europe to eastern Asia. However, there is also a handful of species in the Afrotropics (5). A single species, O. loriae , is known from New Guinea, but the genus is otherwise absent from the Australian and Neotropic regions. For more detailed distribution data of individual species, see Appendix.
Biology. Most current knowledge about orussid biology has been obtained by studying a few of the comparatively common Nearctic ( O. occidentalis , O. sayii ) and Palearctic ( O. abietinus ) species. Extensive reviews of the biology and behaviour of Orussus spp. are found in Cooper (1953), Middlekauff (1983), Ahnlund & Ronquist (2001), and Vilhelmsen et al. (2001). In summary, Orussus spp. appear to be ectoparasites of woodboring insect larvae, preferably Buprestidae (Coleoptera) . Echolocation is apparently paramount for host detection (see Vilhelmsen et al., 2001) and restricts the female wasps to oviposit in dry, barkfree wood (see however, discussion under Orussonia ). The biology of Orussus spp. seems to be representative for the family as a whole, judging from the scarce information available for other species of Orussidae .
Genus Pedicrista Benson, 1935
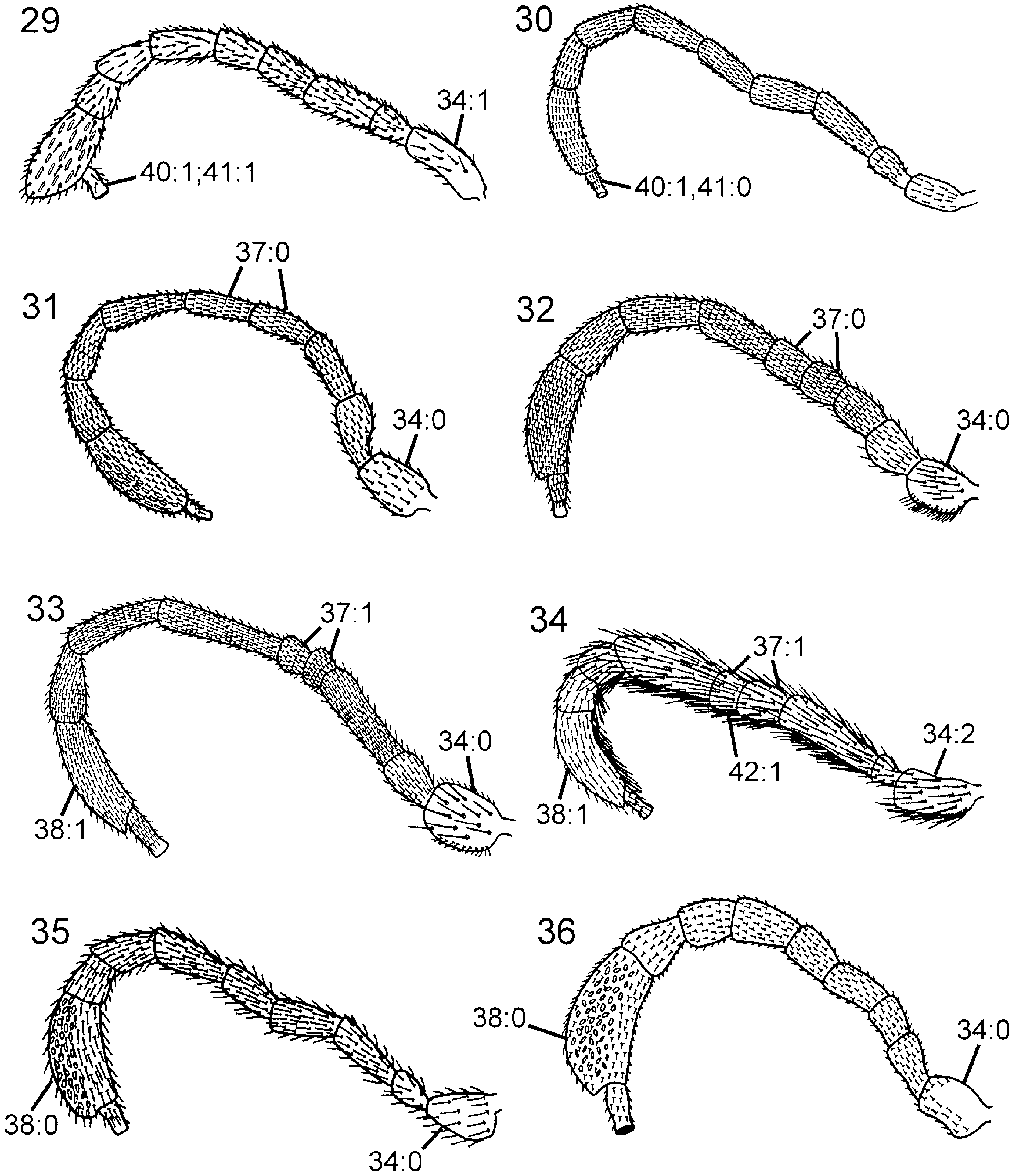
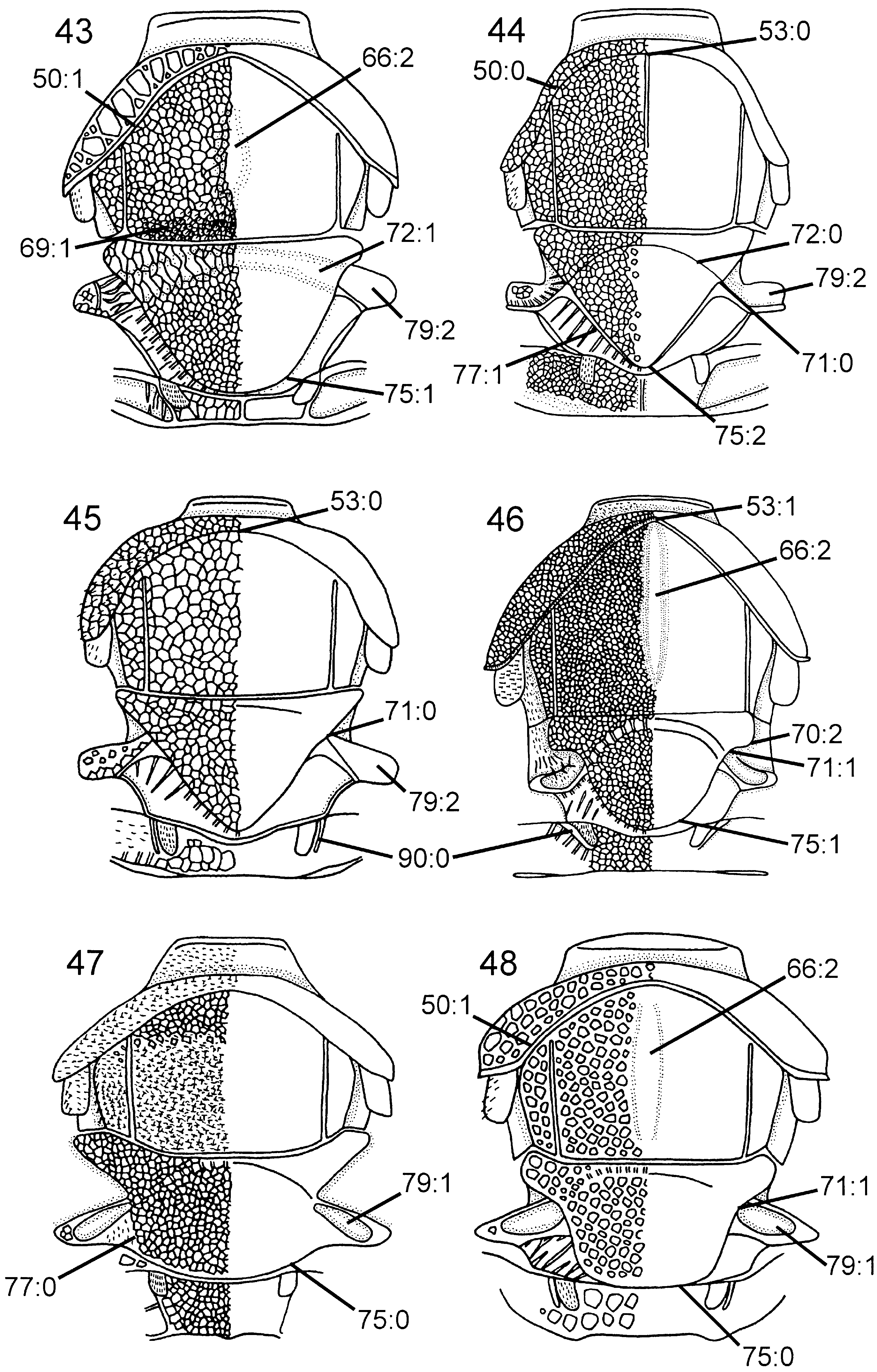
( Figs 5 View Figures 1–6 , 16 View Figures 13–18 , 42 View Figures 37–42 , 53 View Figures 49–54 , 58 View Figures 55–62 , 70 View Figures 63–74 , 102 View Figures 97–105 )
Benson, 1935a: 6. Type species Pedicrista hyalina Benson, 1935a by original designation.
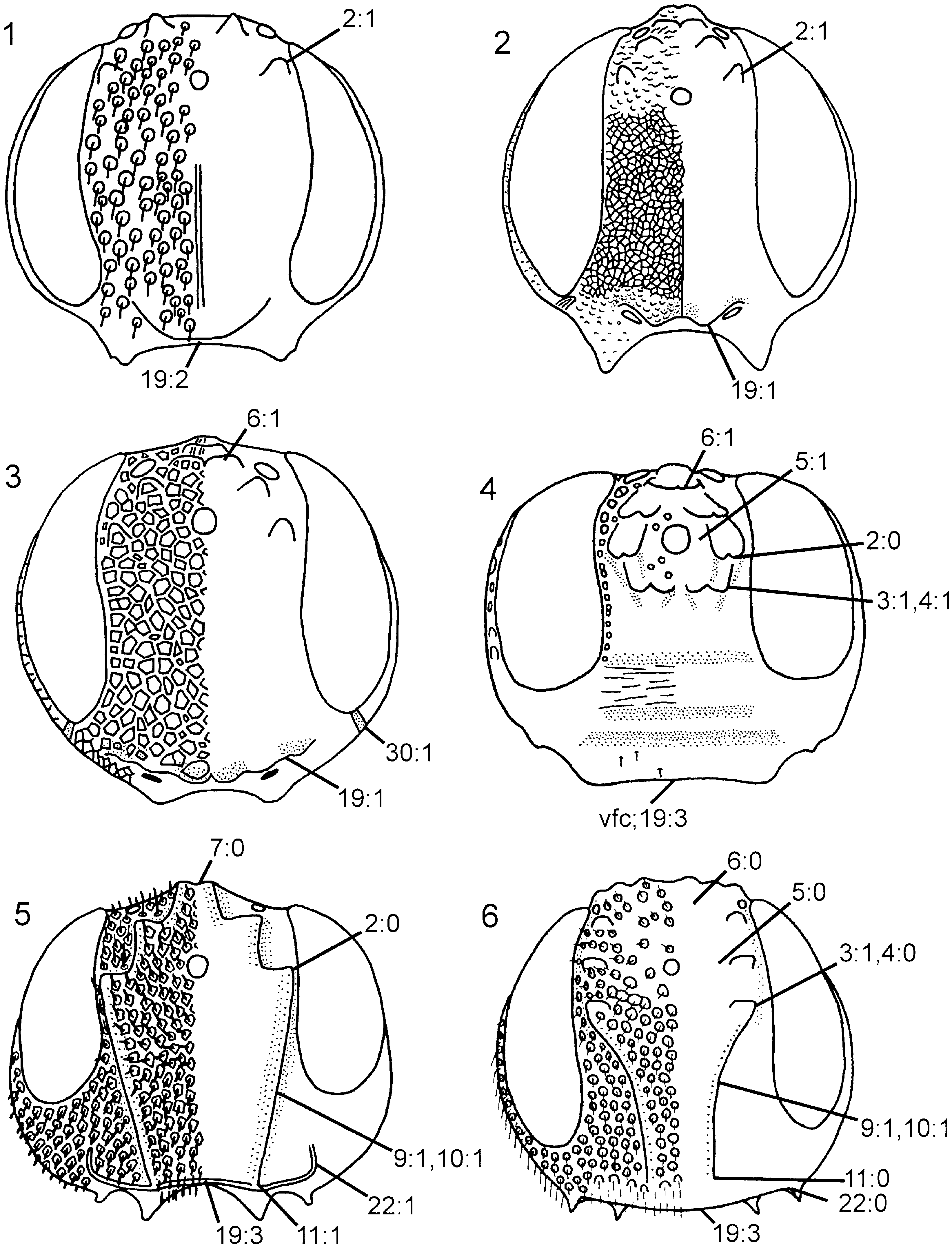
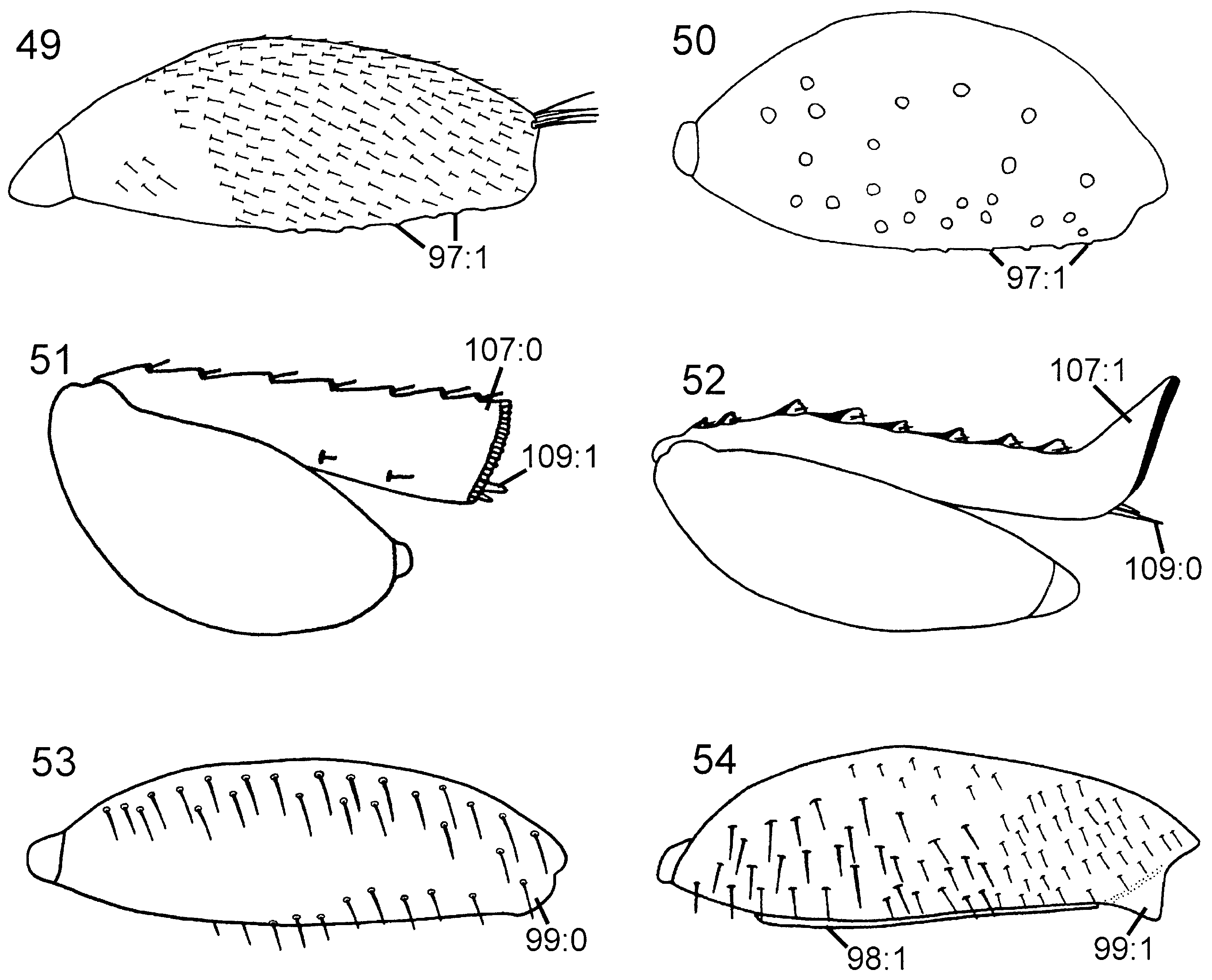
Description. Body and appendages predominantly dark brown, tarsi light brown; light brown markings on femora and proximally on tibiae. Elongate hairs on frons, gena ( Fig. 16 View Figures 13–18 ), dorsal part of pronotum, lateral part of mesopleuron, femora ( Fig. 53 View Figures 49–54 ), tibiae ( Fig. 58 View Figures 55–62 ), and abdomen. Forewing predominantly hyaline ( Fig. 70 View Figures 63–74 ), at most weakly infuscated posterior to pterostigma; pterostigma, veins C and R, and proximal part of vein dark brown, remainder of forewing venation light brown or uncoloured.
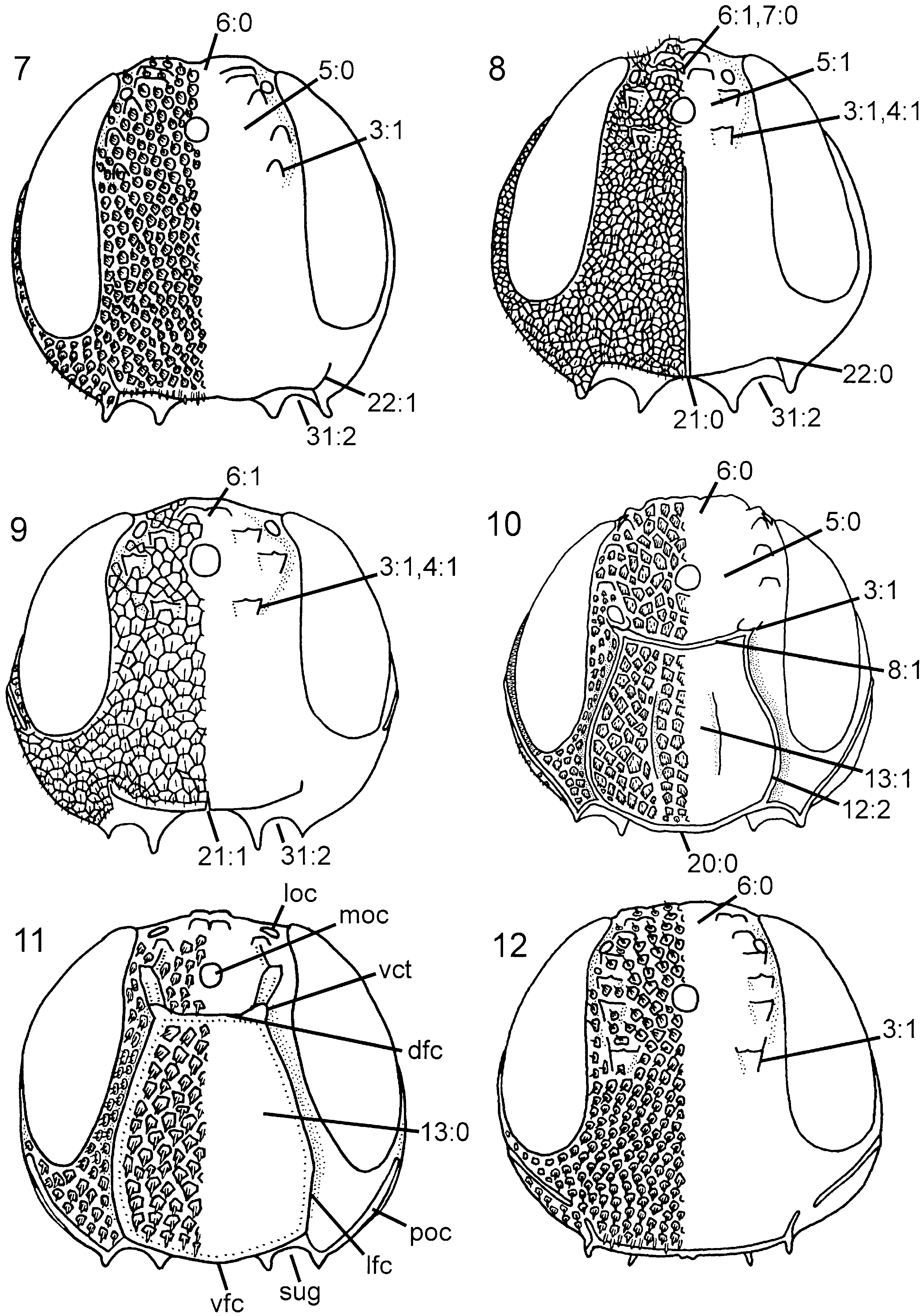
Head. Three pairs of coronal teeth present, dorsalmost pair fused medially; ventral coronal teeth absent ( Fig. 5 View Figures 1–6 ). Lateral ocellus situated well posterior to ocellar corona, separated from median margin of eye by distance equal to its own diameter. Dorsal transverse and lateral longitudinal frontal carinae absent, median longitudinal frontal carinae extend from ventral coronal teeth, converge medially and join ventral transverse frontal carina ( Fig. 5 View Figures 1–6 ); area between median longitudinal frontal carinae distinctly concave. Ventral transverse frontal carina with distinctly upturned lateral ends. Dense pilosity posterior to eye absent, postocular carina well developed, occipital carina absent ( Fig. 16 View Figures 13–18 ). Subantennal groove only weakly developed, delimited laterally by short, low carina. Frons and vertex areolate, occiput reticulate, gena predominantly foveate. Scapus short and subcylindrical. Female antenna with A4 + 5 longer than A6, A9 expanded distally with lateral margin rounded. Labiomaxillary complex reduced, maxillary and labial palps with four and two segments, respectively.
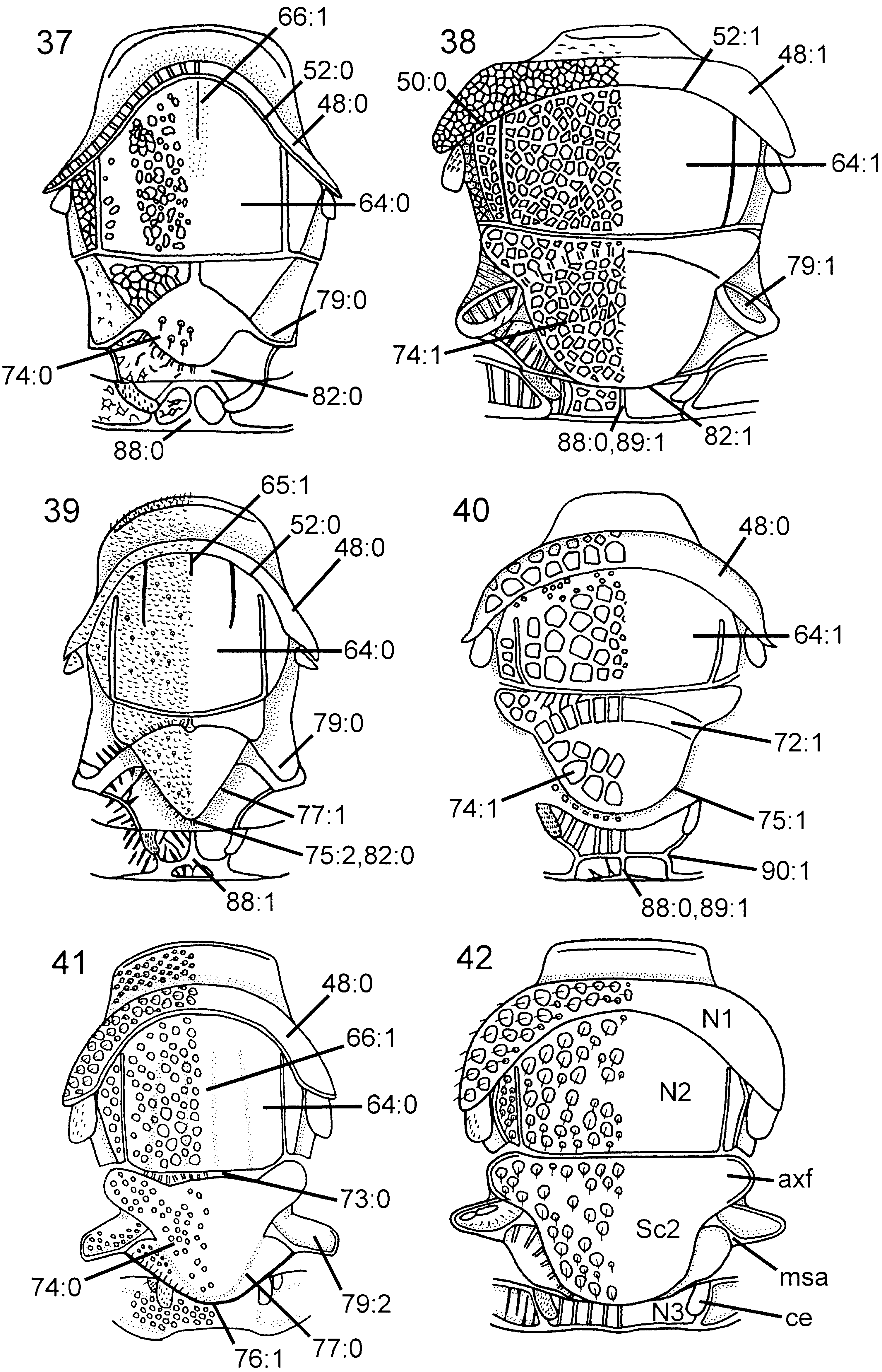
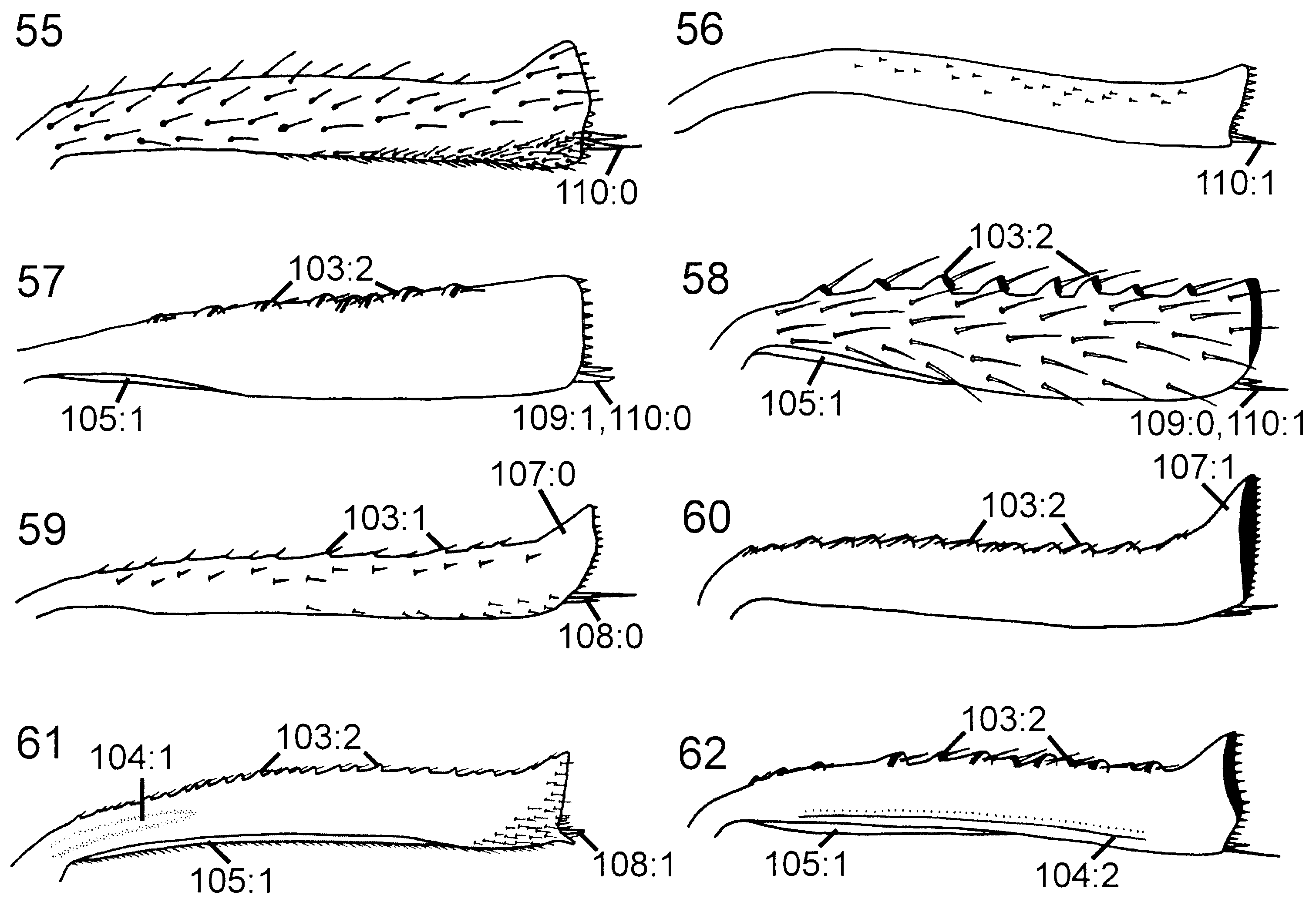
Thorax. Pronotum deeply curved posteriorly, dorsal part longer laterally than medially ( Fig. 42 View Figures 37–42 ). Forecoxa not expanded medially. Mesoscutum in female with slightly raised area medially, mesoscutellar sulcus absent. Mesoscutellum rounded posteriorly, fused with posterior margin of mesonotum, subdividing mesopostnotum medially ( Fig. 42 View Figures 37–42 ). Mesoscutellum not projecting beyond cenchrus, with lateral margin distinct. Mesoscutellar arm distinct and pointed, with shallow depression anteriorly. Mesopleuron with mesosubalar carina well developed. Metascutellum reduced. Dorsal part of pronotum and mesoscutum reticulate to foveate, mesoscutellum foveate with large glabrous interspaces. Mesopleuron reticulate to foveate anterolaterally, glabrous posteriorly and ventrally. Hindcoxa with only scattered hairs laterally. Hindfemur with denticles absent, longitudinal ridge present ventrally, posteroventral corner rounded ( Fig. 53 View Figures 49–54 ). Hindtibia with dorsal pegs well developed, ventral ridge present ( Fig. 58 View Figures 55–62 ). Two apical spurs present on hindtibia, one more than twice as long as the other.
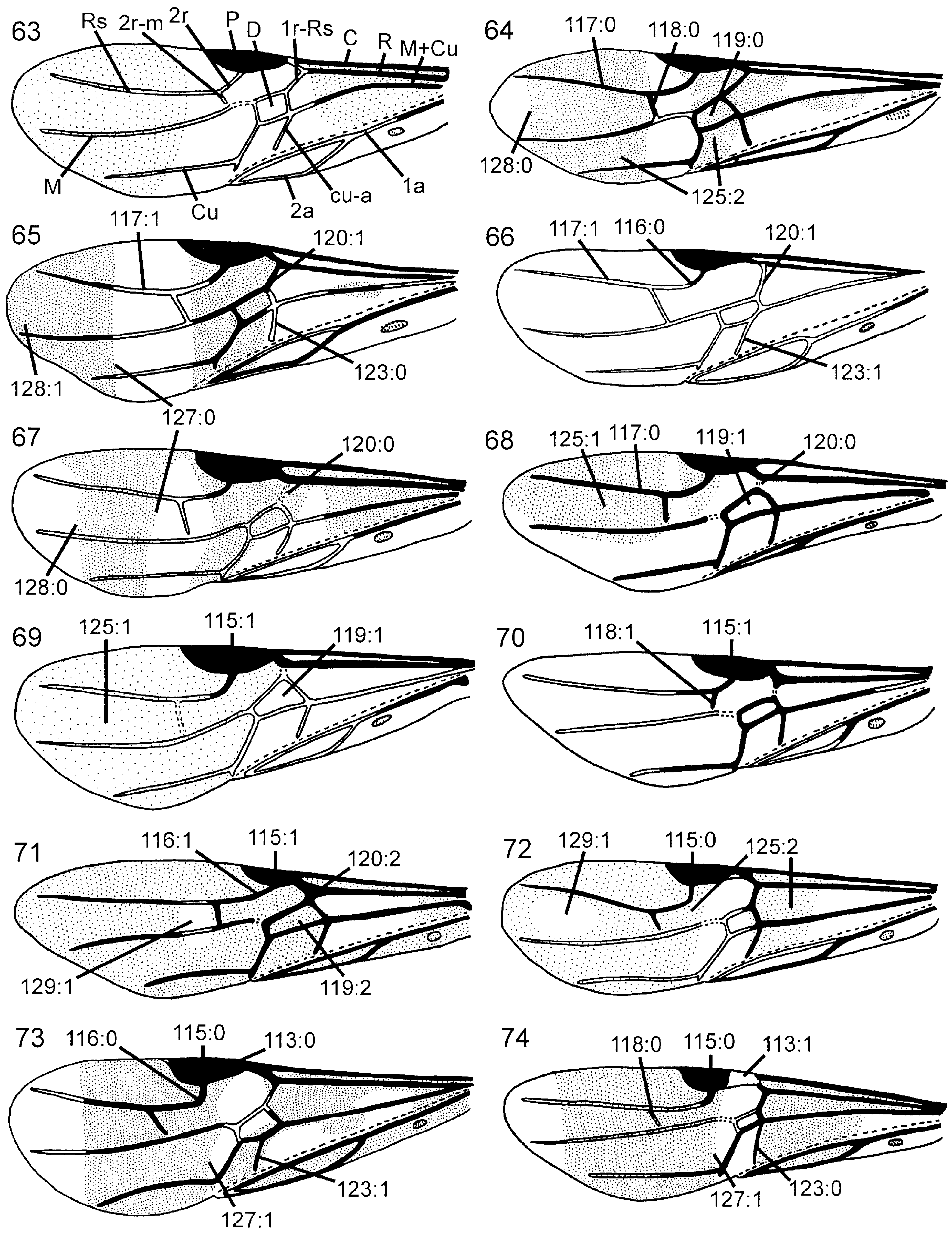
Wings. Vein 2r arises from middle of forewing pterostigma, curving to join vein Rs where 2r-m issues from latter, proximal to distal end of pterostigma ( Fig. 70 View Figures 63–74 ); transition between Rs and 2r distinct. Forewing discal cell of equal length proximally and distally, well separated from vein R. Vein 1r-Rs weakly developed, spectral.
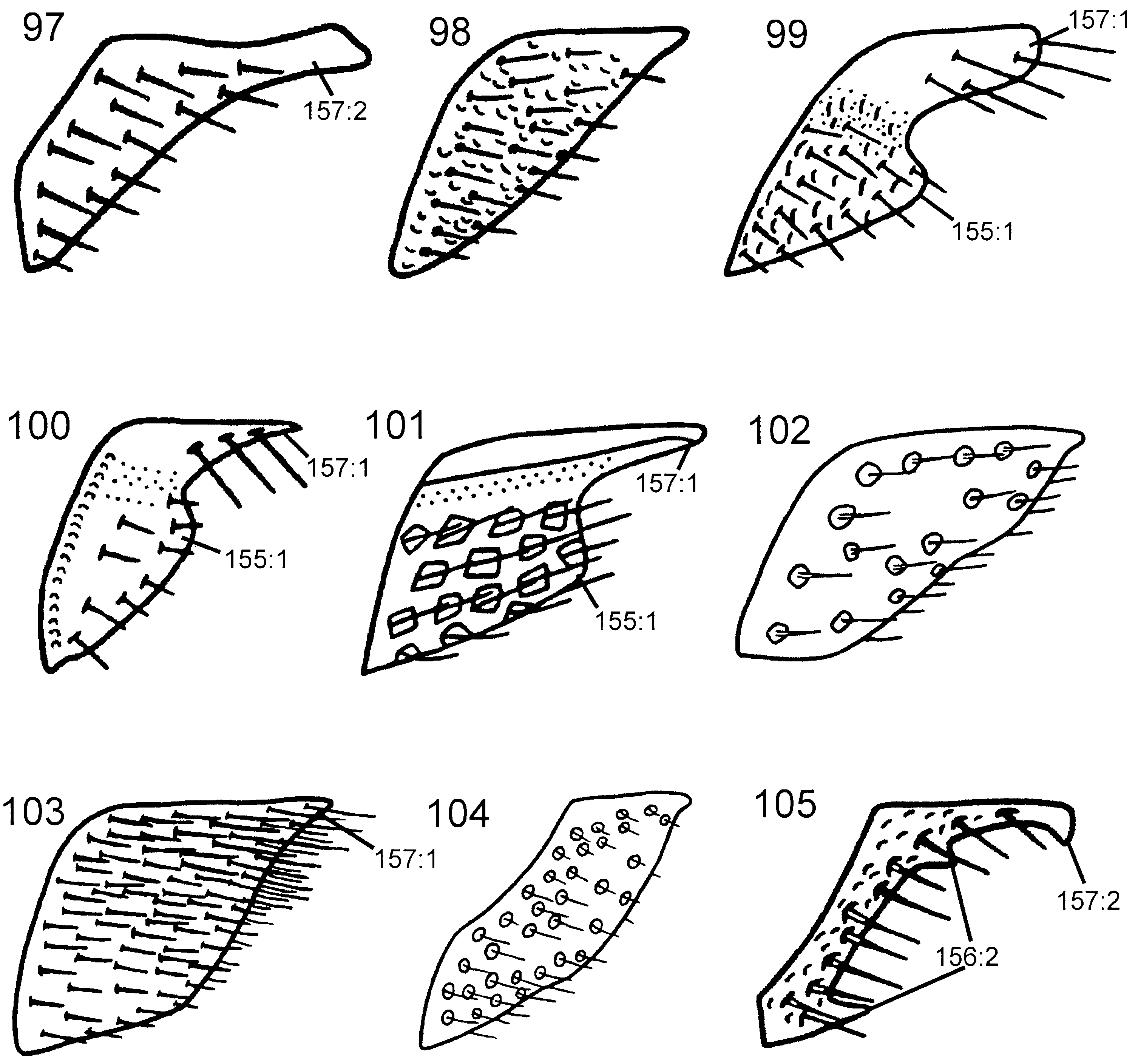
Abdomen. Postspiracular carina on T1 not observed with certainty. T2 with smooth area laterally rectangular and broadly contiguous with antecostal sulcus. Female T8 with small projection posteromedially. Female T9 with longitudinal carina absent, shallow groove along median margin present. Male S9 flat and without cuticular projections ( Fig. 102 View Figures 97–105 ). Anterior abdominal terga (T1-3) areolate to articulate, posterior ones punctate with large glabrous interspaces, with depressed glabrous hindmargins. Abdominal sterna, including male S9, punctate with large glabrous interspaces.
Discussion. Pedicrista is a very distinct taxon; unique features are the position of the lateral ocelli relative to the ocellar corona and the eyes, the frons being concave between the median longitudinal frontal carinae, and the presence of elongate hairs on various body parts (see description). Additional putative autapomorphies are the presence of white markings on the legs (63: 1 i; 111: 1 i), the median subdivision of the mesopostnotum by the posterior margin of the mesoscutellum (82: 1 i), the position of the forewing 2r-m proximal to the distal end of the pterostigma (118: 1 i), and the forewing being almost entirely hyaline in both sexes (125: 0 i; 126: 0 i). The presence of maxillary and labial palps with four and two segments (46: 1), respectively, is unique within the Orussidae ; however, it might be regarded as an intermediate step in a transformation series leading to the even further reduced labiomaxillary complex of Chalinus + Mocsarya , which is probably the sistergroup of Pedicrista . Some features can be interpreted as autapomorphies of Pedicrista in this case also: the absence of an occipital carina (26: 0 i), the subantennal groove with only a weakly developed carina laterally (31: 1 i), and the presence of a distally swollen A 9 in the female antenna (38: 0 i). The first two of these traits are reversals, and all of them could be interpreted as indicating a sistergroup relationship between Pedicrista and Leptorussus ; however, these two genera do not come out together in any of the analyses.
Valid species
Pedicrista hyalina Benson, 1935a . Holotype female (NHML).
Distribution: Malawi, South Africa, Zimbabwe.
Biology. Unknown.
Genus Pseudoryssus Guiglia, 1954
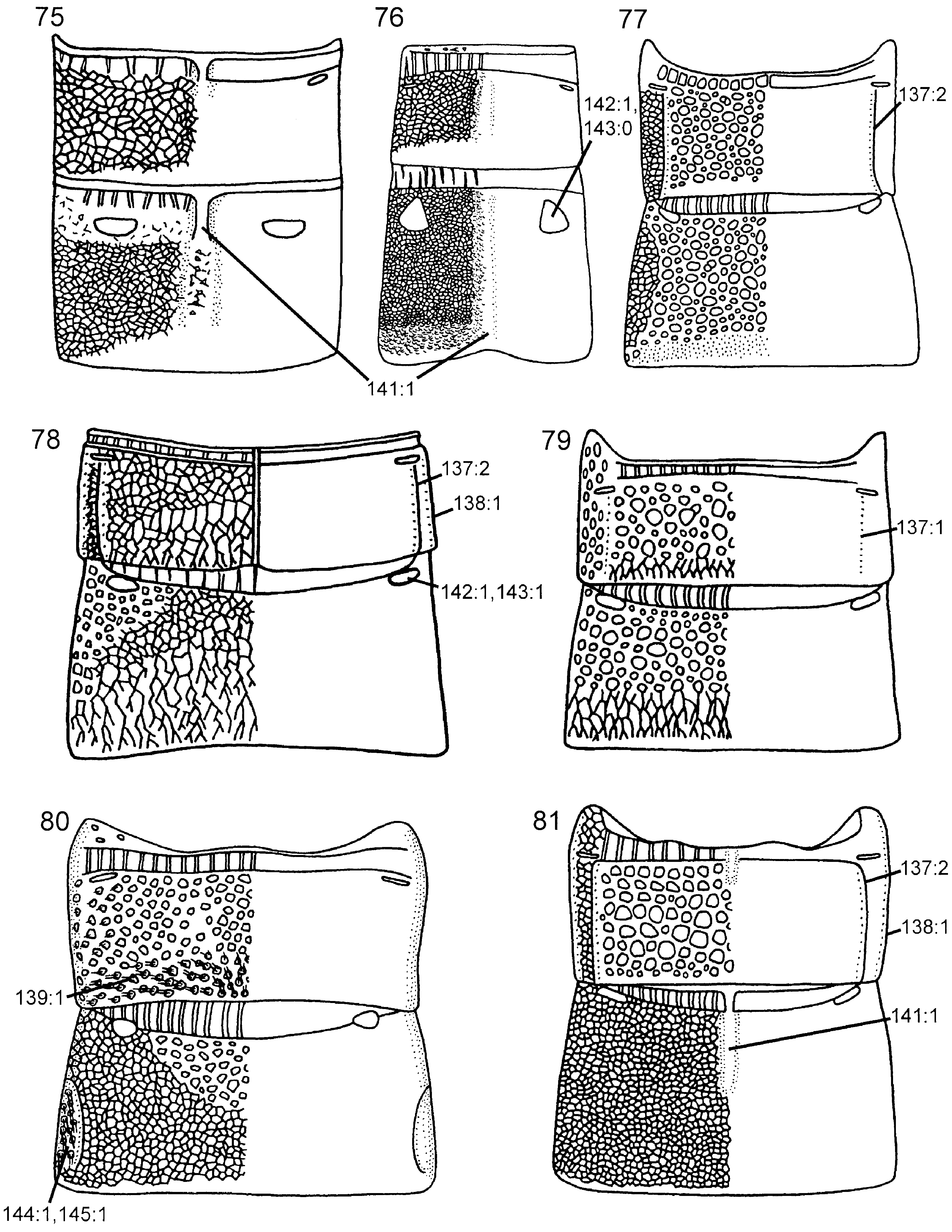
( Figs 9 View Figures 7–12 , 17 View Figures 13–18 , 32 View Figures 29–36 , 45 View Figures 43–48 , 69 View Figures 63–74 , 79 View Figures 75–81 , 83 View Figures 82–87 , 91 View Figures 88–96 , 101 View Figures 97–105 )
Pseudoryssus Guiglia, 1954a: 16 . Type species Orussus henschii Mocsáry, 1910 by original designation ( Guiglia, 1954a View in CoL ).
Description. Body dark brown to black. Appendages mainly dark brown, tarsi light brown. Forewing weakly infuscated from base of pterostigma to apex of wing ( Fig. 69 View Figures 63–74 ); hindwing entirely hyaline. Forewing pterostigma, costal vein, vein R, vein 2r, and proximal part of anal vein dark brown, remaining venation hyaline. Hindwing venation hyaline except for base of subcostal and anal veins.
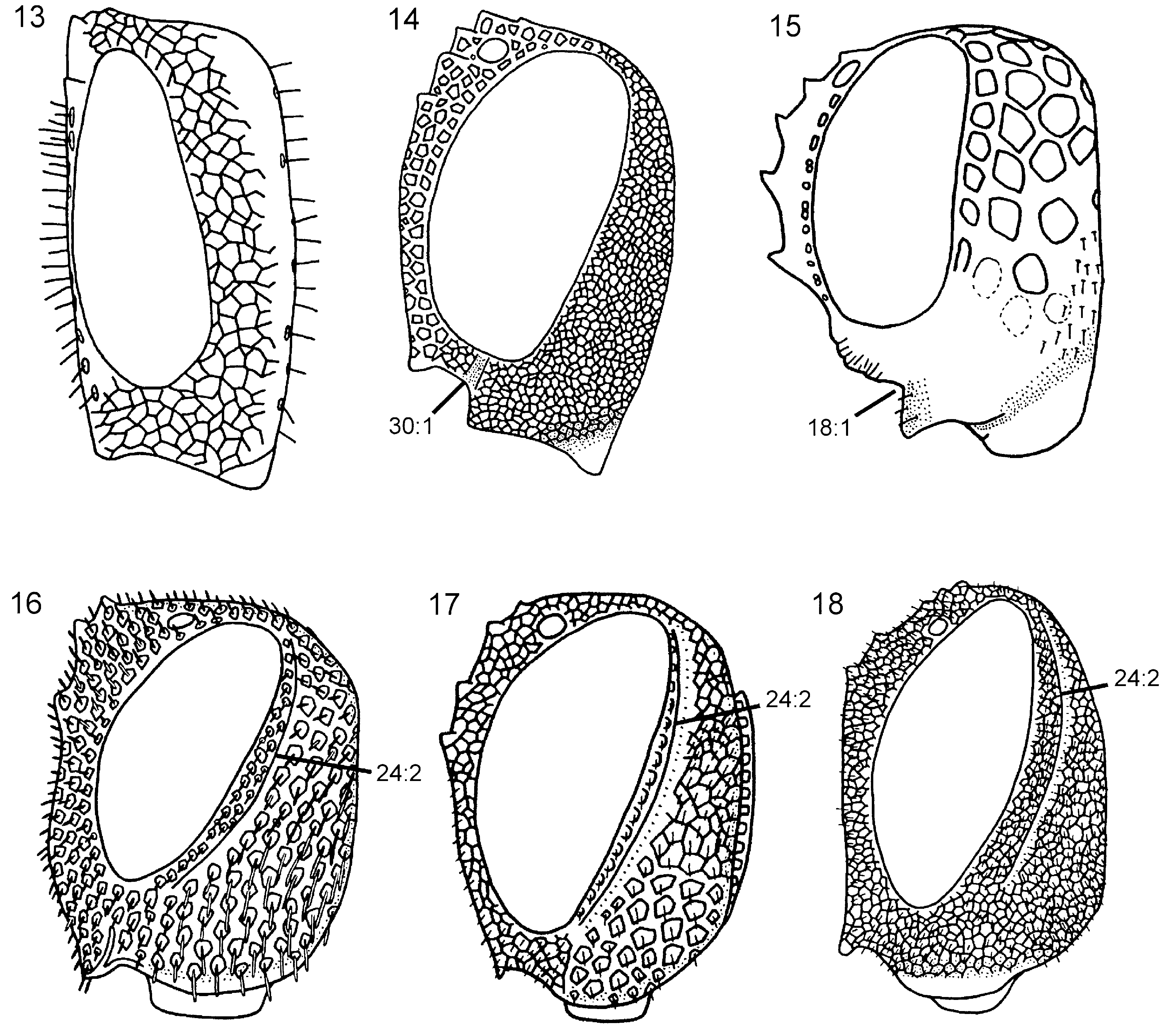
Head. Ocellar corona variable in size, ventral coronal tooth present. Dorsal transverse and longitudinal frontal carinae absent ( Fig. 9 View Figures 7–12 ), ventral transverse frontal carina complete and with notch medially. Pilosity posterior to eye absent, postocular and occipital carinae well developed ( Fig. 17 View Figures 13–18 ). Subantennal groove distinct. Frons and vertex areolate, occiput and gena areolate to reticulate (difficult to see sculpture on P. niehuisorum due to dense pilosity). Scapus short and subcylindrical. Female antenna with A4 + 5 longer than A3 ( Fig. 32 View Figures 29–36 ), A9 not expanded distally and with lateral margin rounded. Labiomaxillary complex fully developed, maxillary and labial palps with five and three segments, respectively.
Thorax. Pronotum variously curved posteriorly and without notch medially in posterior margin, dorsal part longer laterally than medially ( Fig. 45 View Figures 43–48 ). Forecoxa not expanded medially. Mesoscutum in female without median longitudinal ridge, mesoscutellar sulcus indistinct. Mesoscutellum rounded posteriorly, not broadly contiguous with hindmargin of mesonotum, with lateral margin distinct. Mesoscutellar arm distinct and rectangular, without a pit anteriorly ( Fig. 45 View Figures 43–48 ). Mesopleuron with mesosubalar carina well developed. Metascutellum reduced. Dorsal part of pronotum and mesoscutum areolate, mesoscutellum reticulate, mesopleuron reticulate or areolate. Hindcoxa without dense pilosity laterally. Hindfemur without denticles and longitudinal ridge ventrally, posteroventral corner rounded. Hindtibia with one row of ditinct pegs dorsally, no longitudinal ridge ventrally. Hindtibial apical spurs unequal in length, one twice as long as the other.
Wings. Vein 2r arises from middle of forewing pterostigma, with distinct angle where it joins vein Rs. Forewing discal cell longer proximally than distally, not touching vein R anteriorly ( Fig. 69 View Figures 63–74 ); vein 1r-Rs short and spectral. Forewing vein cu-a inserts proximal to discal cell, opposite vein M.
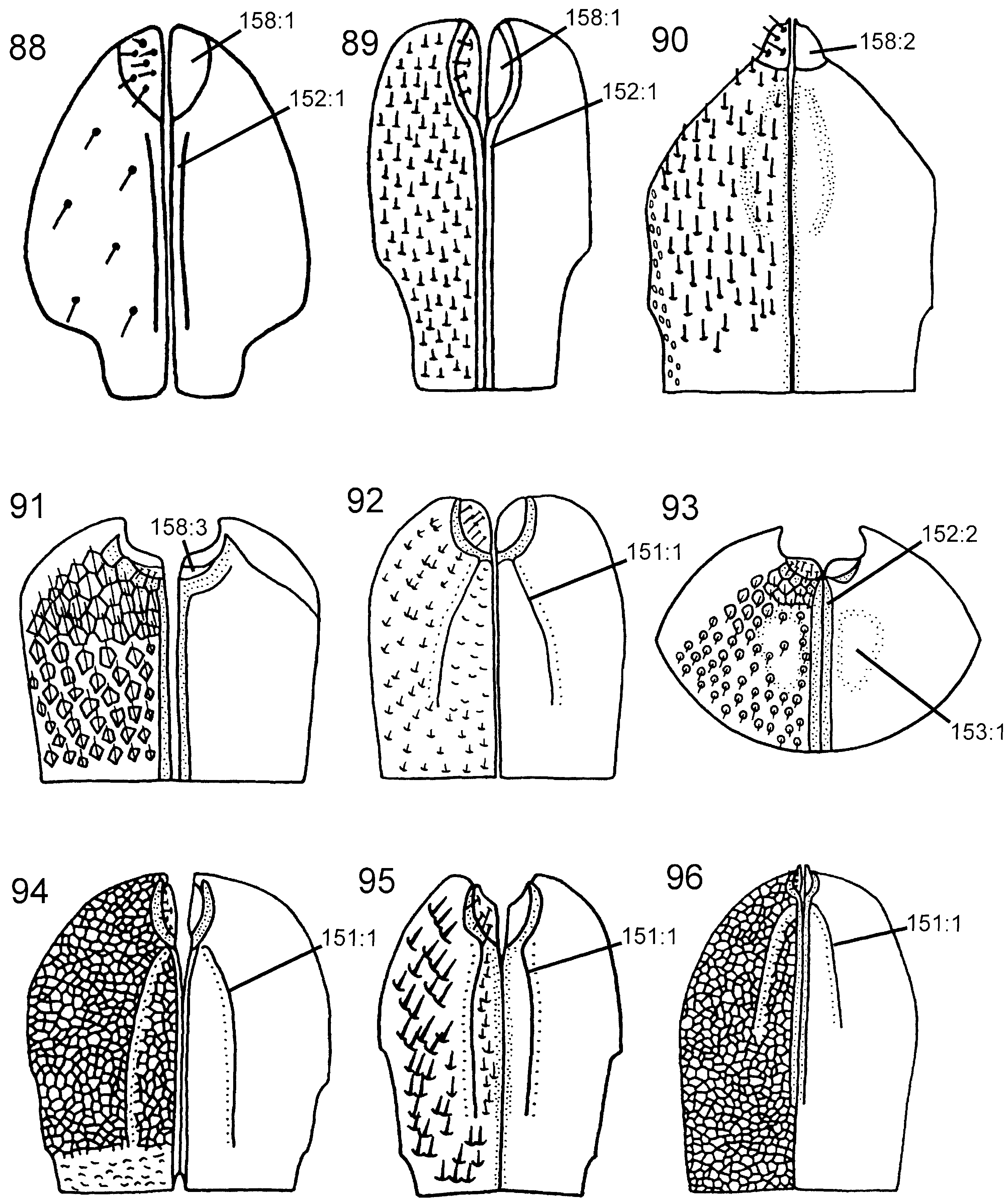
Abdomen. T1 with postspiracular carina weakly developed ( Fig. 79 View Figures 75–81 ). T2 with smooth area laterally rectangular and broadly contiguous with antecostal sulcus. Female T8 with small projection posteromedially ( Fig. 83 View Figures 82–87 ). Female T9 with longitudinal carina absent, distinct groove along median margin present ( Fig. 91 View Figures 88–96 ). Male S9 with posterior margin extended into distinct, tonguelike rim ( Fig. 101 View Figures 97–105 ), without transverse swelling or tubercles. Female T10/cercus rectangular in outline ( Fig. 91 View Figures 88–96 ). Abdominal terga, including female T9, areolate/reticulate-rugose with imbricate interspaces. Abdominal sterna, including male S9, reticulate/punctate-rugose with imbricate interspaces.
Discussion. The monophyly of Pseudoryssus is weakly corroborated, the only putative autapomorphy being the reduced and rectangular female T10/cercus (158: 3 i). Despite this, I consider it preferable to retain it as a separate genus rather than include it in Orussus , which usually comes out as its sistergroup. Pseudoryssus lacks a number of the features present in the ground plan of Orussus (see discussion of that genus), and merging these two genera would weaken the definition of Orussus , as the node connecting them is weakly supported.
Guiglia (1956) View in CoL described the species P. emanuelis from a single female specimen from Cassanola Spinola, Piemonte, Northern Italy. She separated it from the very similar P. henschii by slight differences in the configuration of the ‘anterior margin of the clypeus’ (= ventral transverse frontal carina), the colour, relative size and proportions of the antennal segments, and the shape of the forebasitarsus. No additional specimens have been assigned to P. emanuelis ( Kraus, 1998) . The holotype of P. emanuelis has been compared with that of P. henschii and other specimens of this species. I consider the differences listed by Guiglia (1956) View in CoL between the two nominal taxa insufficient to warrant species status for P. emanuelis , and hence regard it as a junior synonym of P. henschii .
Valid species
P. henschii ( Mocsáry, 1910) . Orussus henschii Mocsáry, 1910 . Holotype female (HNHM).
= P. emanuelis Guiglia, 1956 . syn. nov.
Distribution: Bulgaria, Croatia, Germany, Greece, Iraq, Italy, Morocco, Turkey.
P. niehuisorum Kraus, 1998 . Holotype female (ZSMC).
Distribution: Israel.
Additional material. In addition to the material listed by Kraus (1998: fig. 18) I have seen P. henschii from the following localities: IRAQ, Mosul 1978, C.I.E. leg., 1♀, 2, ‘ On Pistacia’ ( NHML); MOROCCO, Moyen Atlas, Aguelman, Sidi Ali, [no date], S. Benhalima leg., 3♀♀, 1♂, ‘s/[ur] Cèdres’ ( MNHN) .
Biology. The type material was collected from pine wood ( Mocsáry, 1910). Rose (1998) reared P. henschii from a branch of Pyrus communis L. ( Rosaceae ) infested by Clytus arietis (L.) ( Cerambycidae ); he also listed a number of emergence records from different tree species.
Genus Stirocorsia Konow, 1897
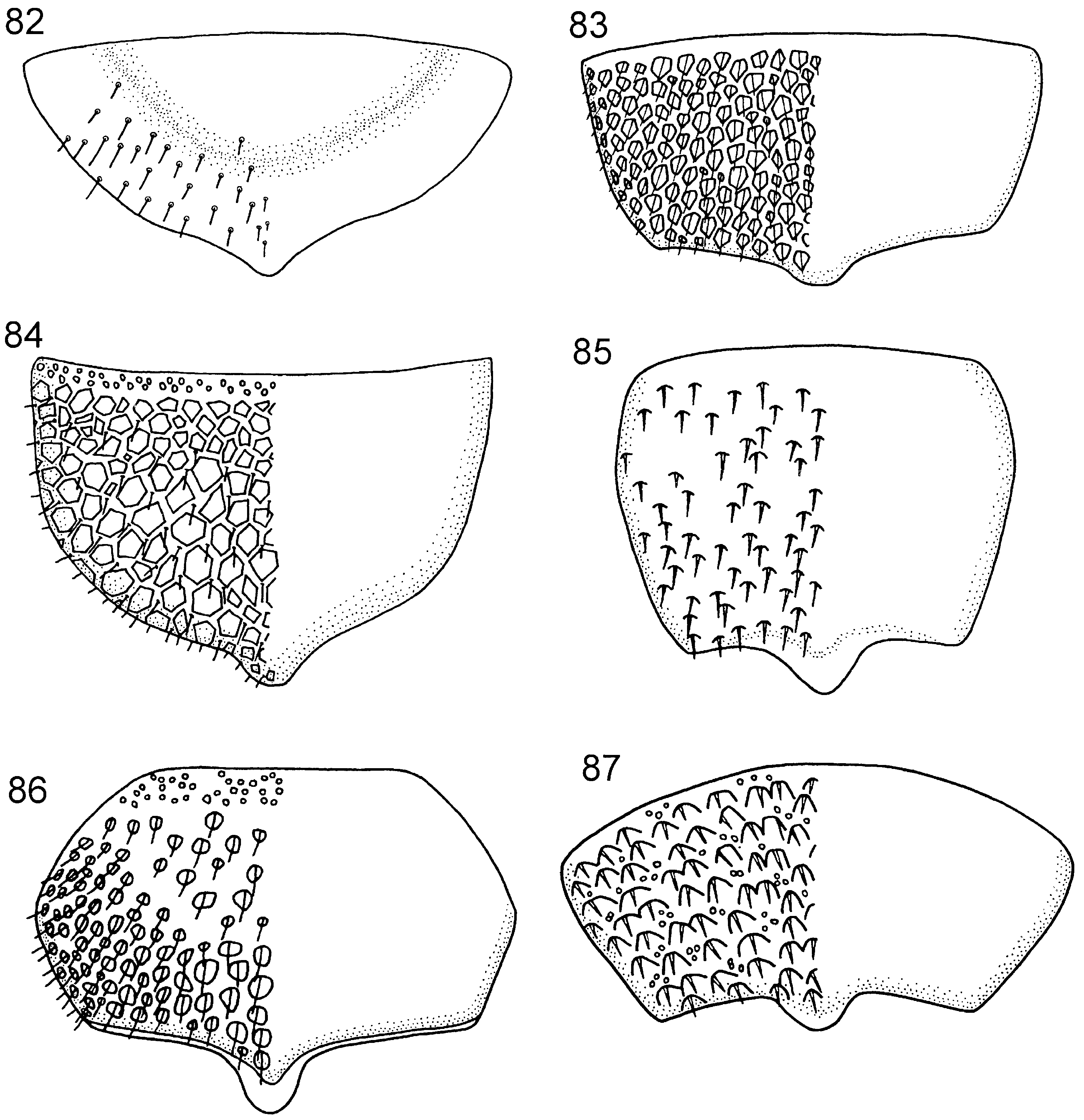
( Figs 24 View Figures 19–24 , 54 View Figures 49–54 , 62 View Figures 55–62 , 86 View Figures 82–87 )
Stirocorsia Konow, 1897b: 372 . Type species Stirocorsia kohli Konow, 1897b , by monotypy.
Description. Body dark brown to black. Appendages variously coloured, at least some femora and tibiae usually reddish brown. Forewing with one or two infuscated bands of various extent; forewing pterostigma usually dark brown, venation predominantly dark brown.
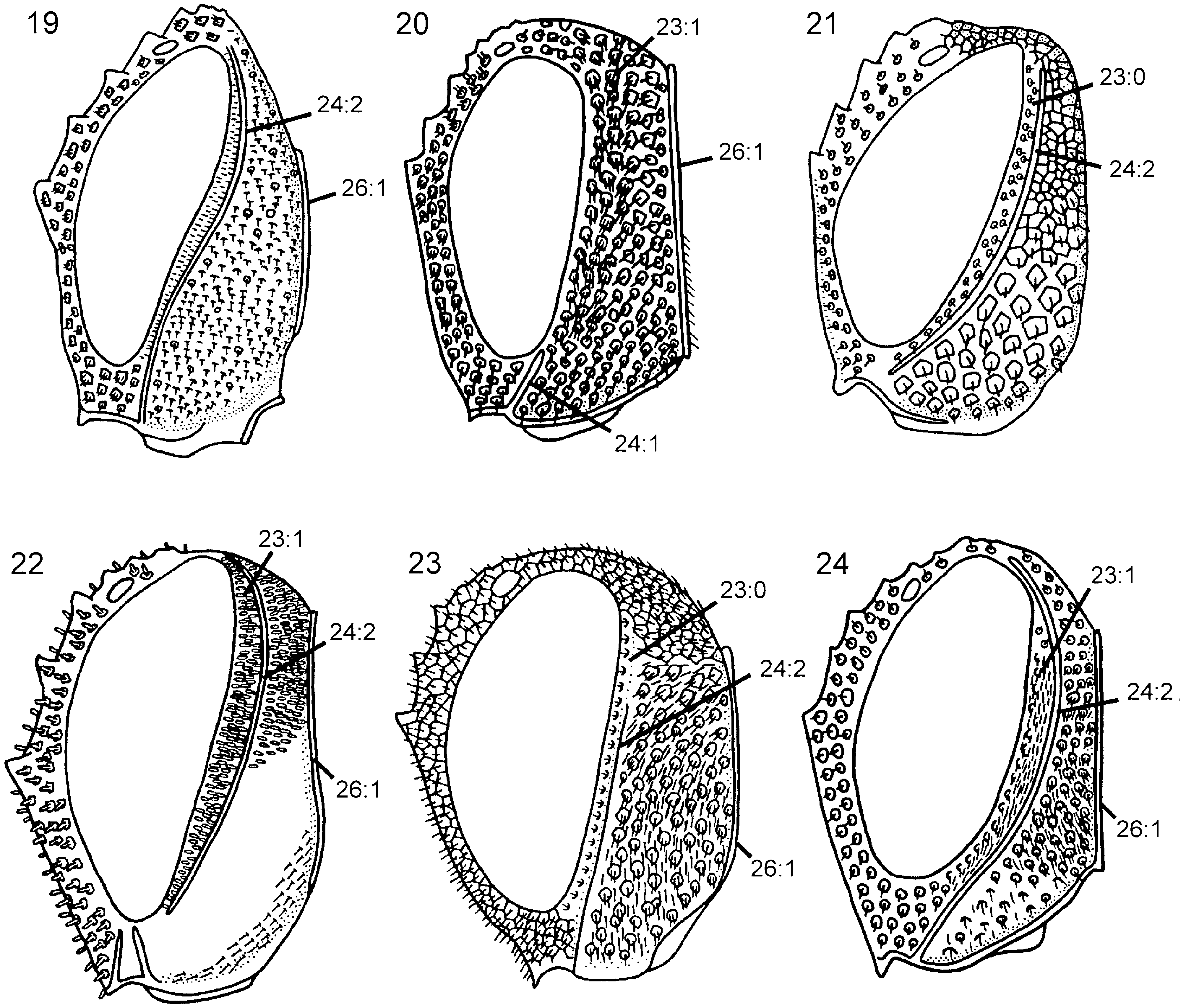
Head. Ocellar corona wide, ventral coronal tooth present; dorsal and ventral transverse frontal carina always present, lateral longitudinal frontal carina usually present, median longitudinal frontal carina always absent; dense pilosity present posterior to eye, postocular and occipital carinae fully developed ( Fig. 24 View Figures 19–24 ); occiput with distinct concavity dorsally; subantennal groove well developed; frons and vertex reticulate to foveate, occiput and gena foveate. Female antenna with combined length of A4 + 5 longer than A6; A9 expanded distally and with carina laterally. Labiomaxillary complex fully developed, maxillary and labial palps with five and three segments, respectively.
Thorax. Pronotum deeply curved posteriorly and with notch medially in hindmargin, dorsal part of pronotum broader laterally than medially; short longitudinal carina present medially on pronotum. Forecoxa not expanded medially. Female mesoscutum medially with well-developed longitudinal carina; mesoscutellar sulcus distinct. Mesoscutellum rounded posteriorly, broadly contiguous with hindmargin of mesonotum; lateral margin of mesoscutellum indistinct, merging with surrounding areas; mesoscutellum not projecting posterior to cenchrus; mesoscutellar arm pointed laterally and with deep pit anteriorly. Mesopleuron with mesosubalar carina present, mesepisternal carina absent. Metascutellum not developed. Pronotum, mesonotum and mesopleuron foveate, metanotum reticulate. Hindcoxa with pilosity laterally well developed. Hindfemur without denticles and usually without longitudinal carina, laterodistal corner triangular ( Fig. 54 View Figures 49–54 ). Hindtibia with two parallel rows (sometimes only developed distally) of distinct pegs dorsally and longitudinal carina ventrally ( Fig. 62 View Figures 55–62 ). Hindtibial apical spurs differ considerably in length.
Wings. Forewing with vein 2r arises from pterostigma about 2/3rds from base of latter, distinct angle present where 2r joins Rs; discal cell of equal length proximally and distally; cu-a inserts proximal to discal cell, opposite vein M.
Abdomen. T1 with postspiracular carina well developed. T2 with smooth lateral area rectangular and continuous with antecostal sulcus anteriorly. Female T8 with well-developed projection posteromedially ( Fig. 86 View Figures 82–87 ). Female T9 with distinct longitudinal carina, delimiting concave area medially. Male S9 obliquely sloping in lateral view, with extended posterior margin terminating in distinct, stubby projection; tubercles small, but distinct. Abdominal tergal sculpture variable, usually reticulate to foveate on anterior terga (T1-2), foveate to punctate on posterior terga (T3-8); female T9 rugose-reticulate. Abdominal sterna foveate to punctate, male S9 reticulate-rugose.
Discussion. The monophyly of Stirocorsia is rather weakly supported, the only good autapomorphy being the presence of a short longitudinal carina medially on the dorsal part of the pronotum (49: 1; see Vilhelmsen & Smith, 2002: fig. 18); this is a unique feature, but still has to be confirmed for S. apicalis , which was not available for study. Benson (1935a) regarded Stirocorsia as a synonym of Ophrynopus ; however, Benson (1938b) reestablished Stirocorsia after revising which species should be included. Benson (1938b) has been followed here because even though Stirocorsia resembles Ophrynopus in many traits, it does not come out as the sistergroup of the latter in all analyses (see discussion of generic relationships below).
Valid species
S. apicalis Togashi, 2000 . Holotype male (NSMT).
Distribution: Japan ( Vilhelmsen & Smith, 2002: fig. 42).
S. kohli Konow, 1897b . Holotype female (NHMW).
= S. cariniceps Cameron, 1905 . Holotype female (NHML). Synonymized by Benson (1935b).
= O. philippinensis Guiglia, 1937e . Holotype female (EITH). Synonymized by Vilhelmsen & Smith (2002).
Distribution: Indonesia, Laos, Malaysia, Philippines ( Vilhelmsen & Smith, 2002: fig. 41).
S. maculipennis ( Smith, 1859) . Holotype female ( OXUM).
= O. trifasciata Cameron, 1906 . Holotype female (ZOMU). Synonymy reestablished by Vilhelmsen & Smith (2002).
= O. rossi Yasumatsu, 1952 . Holotype female (KUEC). Synonymized by Vilhelmsen & Smith (2002).
Distribution: Indonesia, Papua New Guinea ( Vilhelmsen & Smith, 2002: fig. 41).
S. tosensis ( Tosawa & Sugihara, 1934) . Holotype female, depository unknown; according to Guiglia (1966) View in CoL ‘Tipo in Coll. Tosawa ( Osaka) [private collection]’.
Distribution: Japan ( Vilhelmsen & Smith, 2002: fig. 42).
Biology. S. tosensis has been collected from dead wood of Quercus ( Fagaceae ( Guiglia, 1966) .
| NHML |
Natural History Museum, Tripoli |
| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Orussus sayii
| Vilhelmsen, Lars 2003 |
Pseudoryssus
| Guiglia D 1954: 16 |
Stirocorsia
| Konow FW 1897: 372 |