Ophrynopus
Orussonia
Orussella
Orussobaius
Leptorussus
Pseudoryssus
Orussus
Pedicrista
Mocsarya
Chalinus
Ophrynon
Argentophrynopus
Kulcania
Guiglia chiliensis
Guiglia
coracina
Guiglia sericata
Guiglia bombycinis
Guiglia
rubricata
Guiglia schauinslandi
Guiglia rubicunda
Ophrella
Stirocorsia
Ophrynopus
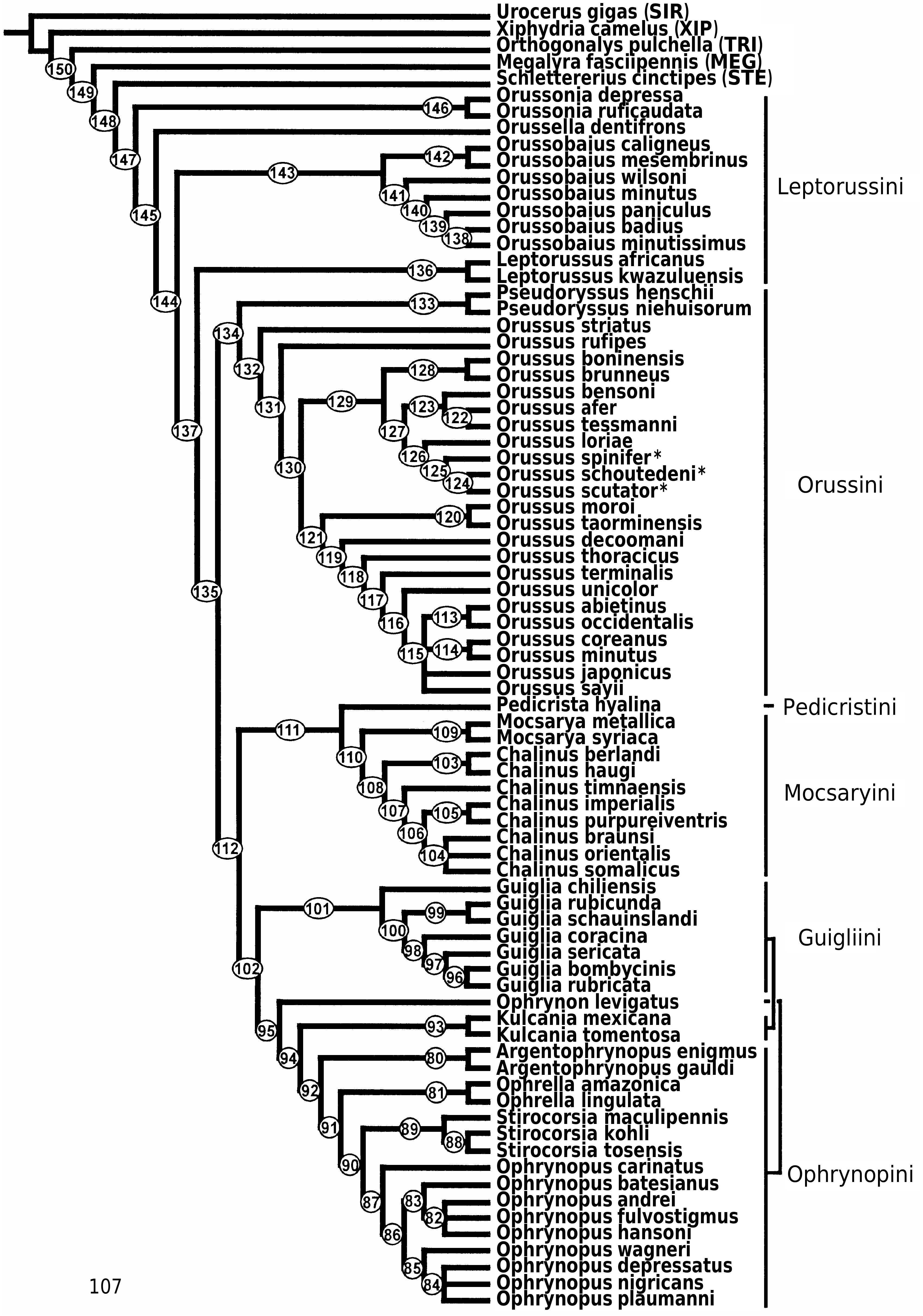
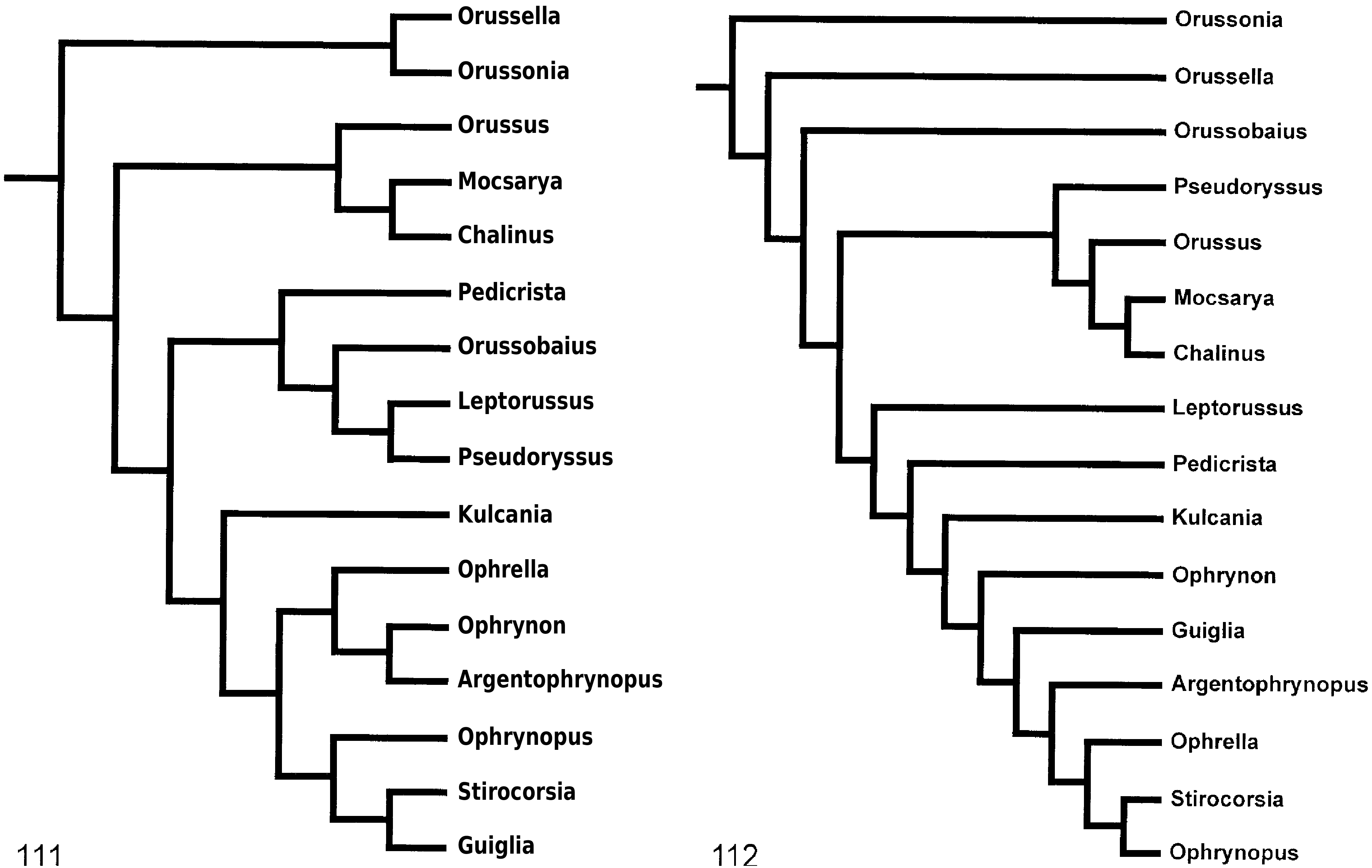
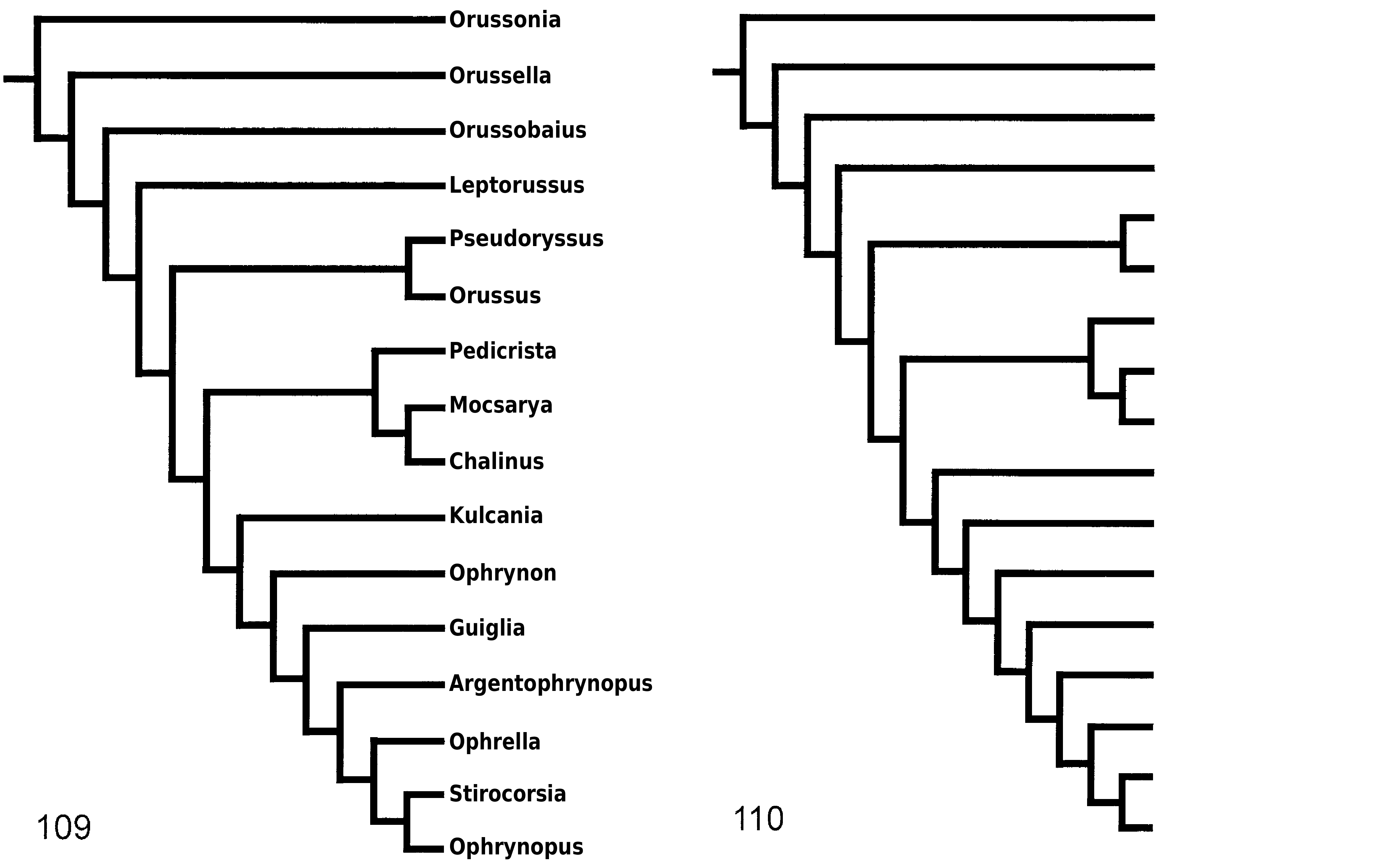
margins of the mesoscutellum (77: 2 i); sistergroup of the ‘ophrynopine’ genera (conc3; k = 2–3 ( Fig. 112
View Figures 111–112
), supported by the presence of a ventral carina on the hindtibia (105: 1 i); sistergroup of (
Orussus
+ (
Chalinus
+
Mocsarya
)) +
Pseudoryssus
(k = 1; Fig. 113
View Figures 113–114
), supported by the forewing vein 1r-Rs being spectral (120: 0 c). None of these alternatives seem well corroborated in comparison with the preferred topology.
Pseudoryssus
is usually placed as the sistergroup of
Orussus
( Figs 106
View Figure 106
, 107
View Figure 107
, 109, 110
View Figures 109–110
, 114
View Figures 113–114
, 115
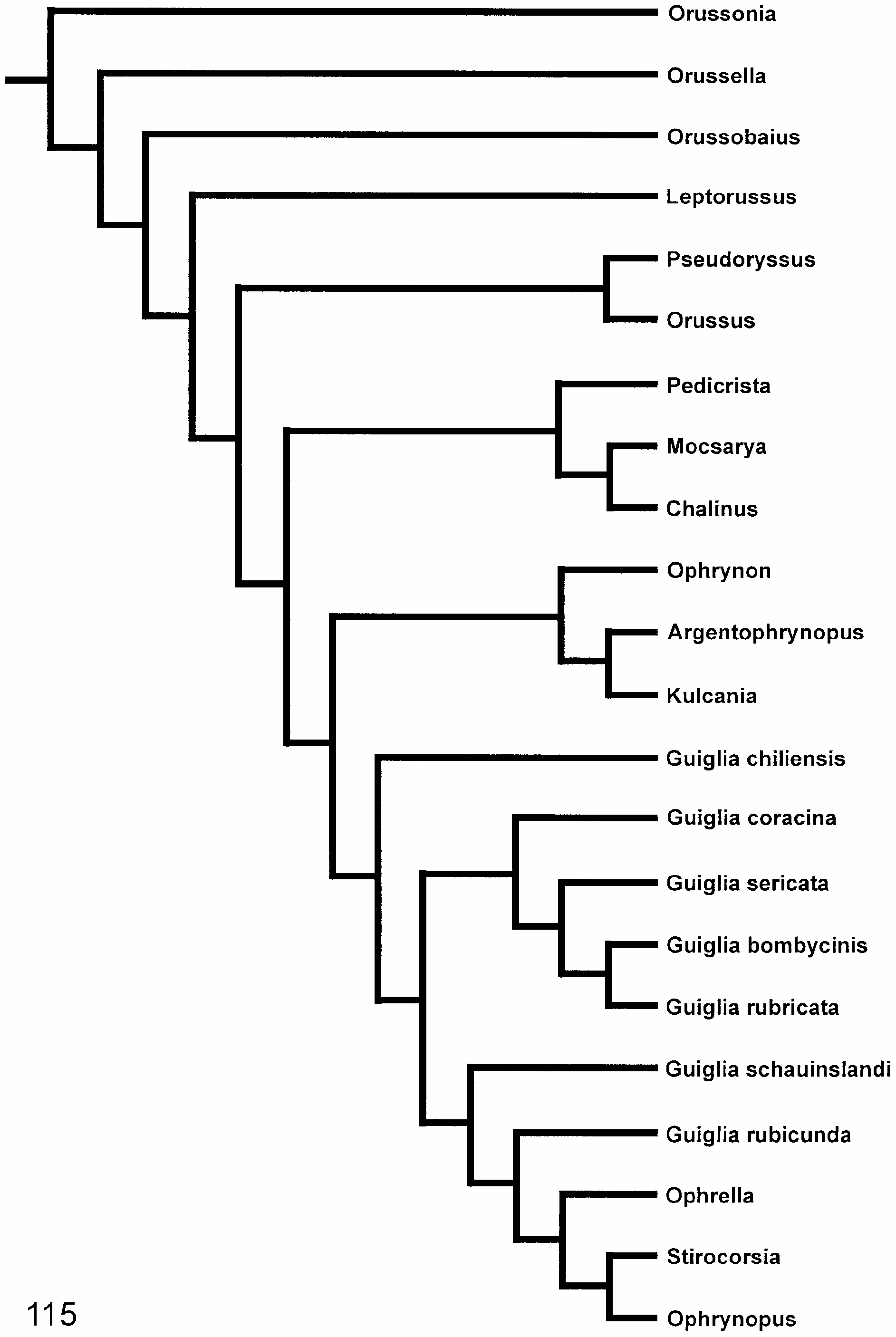
View Figure 115
). The support is rather weak, putative synapomorphies being the mesoscutellar arm having a raised area without a pit anteriorly (79: 2 i), the forewing discal cell being rhomboid (119: 1 c,i,r), and, under some weighting schemes (conc5–6), the forewing vein 1r-Rs being spectral (120: 0 i).
Pseudoryssus
rarely comes out as the sistergroup of
Leptorussus
(conc1–2; Fig. 111
View Figures 111–112
, see also above), but occasionally as sistergroup to (
Chalinus
+
Mocsarya
) +
Orussus
(conc3; k = 1–3 ( Figs 112
View Figures 111–112
, 113
View Figures 113–114
). The latter topology is supported by the same characters as the preferred one, the differences being caused by the conflict between the characters supporting (
Chalinus
+
Mocsarya
) +
Orussus
(see above) and ((
Chalinus
+
Mocsarya
) +
Pedicrista
) + ‘ophrynopine’ genera.
The ‘ophrynopine’ genera (
Argentophrynopus
,
Guiglia
,
Kulcania
,
Ophrella
,
Ophrynon
,
Ophrynopus
and
Stirocorsia
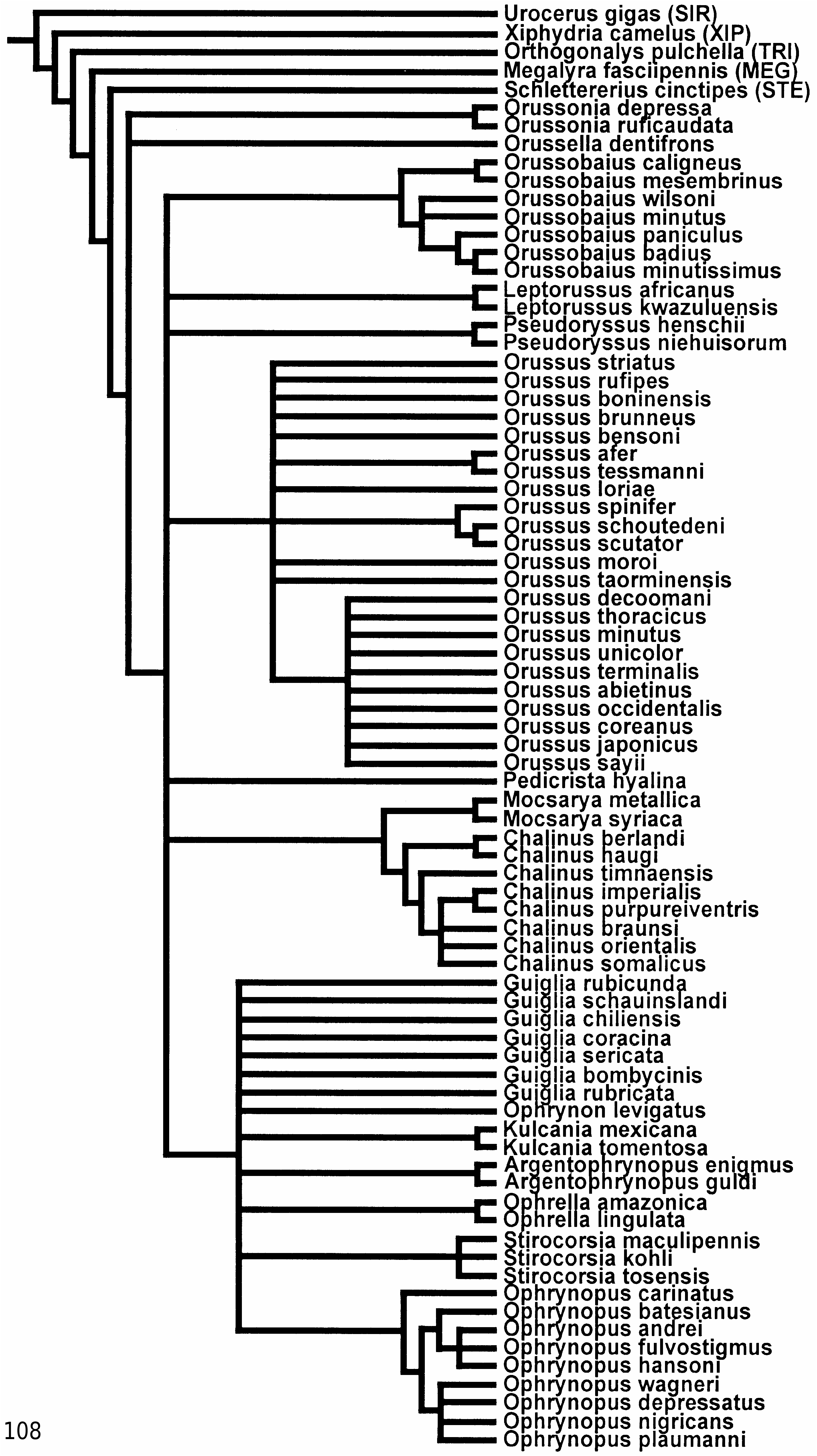
) constitute a clade that is retrieved under all weighting conditions ( Fig. 108
View Figure 108
). The most convincing synapomorphies are the distal position of the forewing vein 2r relative to the pterostigma (115: 0 i), the presence of a longitudinal carina laterally on the female 9th abdominal tergum (151: 1 i), and the presence of tubercles on the male 9th abdominal sternum (156: 1 c,i). The sistergroup of the ‘ophrynopine’ genera usually is (
Chalinus
+
Mocsarya
) +
Pedicrista
( Figs 106
View Figure 106
, 107
View Figure 107
, 109, 110
View Figures 109–110
, 114
View Figures 113–114
, 115
View Figure 115
); this topology is rather weakly supported, the strongest support coming from having a carina developed ventrally on the hindtibia (105: 1 i,r) and having a transverse swelling absent from the male 9th abdominal sternum (155: 0 i,r). Rarely occurring alternative sistergroups to the ‘ophrynopine’ genera are all other genera except
Orussonia
,
Orussella
and
Orussobaius
(k = 1; Fig. 113
View Figures 113–114
), being supported by the characters mentioned above for the clade comprising all genera of
Orussidae
except the three latter, and ((
Leptorussus
+
Pseudoryssus
) +
Orussobaius
) +
Pedicrista
(conc1–2; Fig. 111
View Figures 111–112
), a relationship supported mainly by the presence of a carina ventrally on the hindtibia (105: 1 i,r). The support for
Pedicrista
alone as the sistergroup of the ‘ophrynopine’ genera (conc3; k = 2–3 ( Fig. 112
View Figures 111–112
) has already been discussed.
The relationships between the ‘ophrynopine’ genera are weakly supported and changes substantially depend on which weighting scheme is implemented.
Guiglia
is placed as sistergroup of the remaining genera under many weighting conditions, including the preferred topology (conc = 4; k = 4–12 ( Figs 107
View Figure 107
, 114
View Figures 113–114
). The ‘ophrynopine’ genera excluding
Guiglia
are supported by having the 9th female antennomere swollen subapically (38: 0 i) and a carina present laterally (39: 1 i). An alternative topology (CI; RC; RI; conc1–3, 5–6; k = 1–3 ( Figs 109–113
View Figures 109–110
View Figures 111–112
View Figures 113–114
) has
Kulcania
as the basalmost ‘ophrynopine’ genus; the other genera are supported by the presence of a distinct triangular projection on the posteroventral corner of the hindfemur (99: 1). A third possibility is (
Argentophrynopus
+
Kulcania
) +
Ophrynon
(k = 13 +; Fig. 115
View Figure 115
) as the sistergroup of the remaining ‘ophrynopine’ genera; the former clade is supported by the antennal characters listed above as support for the ‘ophrynopine’ genera exclusive of
Guiglia
, the latter clade by the absence of a distinct carina posteriorly on the mesoscutellar arm (80: 0 r).
Argentophrynopus
is the most recently described genus in the
Orussidae ( Vilhelmsen & Smith, 2002)
View in CoL
. A survey of its changing position within the ‘ophrynopine’ clade under different weighting conditions corroborates that it should not be merged with any of the other genera.
Argentophrynopus
turns up as the sistergroup of ((
Ophrynopus
+
Stirocorsia
) +
Ophrella
) +
Guiglia rubicunda
(RI; Fig. 110
View Figures 109–110
), of (
Ophrynopus
+
Stirocorsia
) +
Ophrella
(CI, RC; conc3, 5–6; k = 3, 6–12 ( Figs 107
View Figure 107
, 109
View Figures 109–110
, 112
View Figures 111–112
), of
Ophrynon
(conc1–2; k = 1–2 ( Figs 111
View Figures 111–112
, 113
View Figures 113–114
), and of
Kulcania
(conc4; k = 4–5, 13 + ( Figs 114
View Figures 113–114
, 115
View Figure 115
). The topology ((
Ophrynopus
+
Stirocorsia
) +
Ophrella
) +
Argentophrynopus
occurs in the preferred tree ( Fig. 107
View Figure 107
) and is supported by the presence of an indistinct lateral margin on the mesoscutellum (77: 0 i); the sistergroup relationship between
Argentophrynopus
and
Ophrynon
is supported by the presence of a carina ventrally on the forefemur (56: 1 i). Additional support for these and for the other possible placements of
Argentophrynopus
is weak, being provided by characters that are higly variable even within the ‘ophrynopine’ clade.
Guiglia
View in CoL
is the only genus in the
Orussidae
View in CoL
that is not retrieved under all weighting conditions. In most cases (CI; RC; conc1–6; k = 1–12 ( Figs 107
View Figure 107
, 109
View Figures 109–110
, 111–114
View Figures 111–112
View Figures 113–114
) the monophyly of the genus is corroborated, although the internal relationships vary; however,
Guiglia
View in CoL
occasionally comes out as paraphyletic relative to (
Ophrynopus
+
Stirocorsia
) +
Ophrella
(k = 13 +; Fig. 115
View Figure 115
) or rarely as polyphyletic (RI), displaying the topology ((((
Ophrynopus
+
Stirocorsia
) +
Ophrella
) +
G. rubicunda
) +
Argentophrynopus
) + remaining
Guiglia spp.
( Fig. 110
View Figures 109–110
). Character support for the monophyly of
Guiglia
View in CoL
is weak, but so are the characters that corroborate the para- and polyphyletic topologies. I consider it most prudent to retain
Guiglia
View in CoL
as a valid genus until more evidence can be acquired. The problems with retrieving
Guiglia
View in CoL
as monophyletic may be caused by missing data for
G. rubicunda
. This species is only known from a single male specimen, and thus it could not be scored for the characters specific for the female sex. The examination of female
G. rubicunda
specimens might provide additional support for the inclusion of this species in
Guiglia
View in CoL
and perhaps strengthen the case for the monophyly of the genus.
The clade (
Ophrynopus
+
Stirocorsia
) +
Ophrella
is retrieved under most weighting conditions ( Figs 106
View Figure 106
, 107
View Figure 107
, 109, 110
View Figures 109–110
, 112
View Figures 111–112
, 114
View Figures 113–114
, 115
View Figure 115
); it is supported by the unique trait of having the mesoscutellum rounded posteriorly and lying parallel with the anterior margin of the metanotum (75: 0). The sistergroup relationship between
Ophrynopus
and
Stirocorsia
also comes out in most of the analyses; this is supported by the dorsally extended ocellar corona (6: 0 i) and by the presence of a well developed subspiracular carina (138: 1 i). An additional putative synapomorphy for these two genera is the presence of an occipital concavity dorsally on the head (28: 1 i); however, this trait is absent from
Ophrynopus carinatus
. In a few instances,
Stirocorsia
is the sistergroup of
Guiglia
(conc1–2; k = 1–2), in which case
Ophrynopus
is either most closely related to the former two genera combined (conc1; Fig. 111
View Figures 111–112
) or to
Ophrella
(conc2; k = 1–2 ( Fig. 113
View Figures 113–114
). The
Ophrynopus
+
Ophrella
clade is corroborated by the presence of a lateral metanotal carina (90: 1 i), but otherwise support for these relationships is weak.