
Rowlandius linsduarteae, Santos, Adalberto J., Dias, Sidclay C., Brescovit, Antonio D. & Santos, Pollyanna P., 2008
|
publication ID |
https://doi.org/10.5281/zenodo.183403 |
|
DOI |
https://doi.org/10.5281/zenodo.6229809 |
|
persistent identifier |
https://treatment.plazi.org/id/03D387F0-FFAA-FFC6-FF50-F8ABFCF6FB4A |
|
treatment provided by |
Plazi |
|
scientific name |
Rowlandius linsduarteae |
| status |
sp. nov. |
Rowlandius linsduarteae View in CoL sp. nov.
Figures 1–6 View FIGURES 1 – 6 , 9 View FIGURE 9
Rowlandius new species. Santos & Pinto-da-Rocha 2008: fig. 15.
Type material. Holotype male from Área de Proteção Permanente Mata do Buraquinho ( 07°06'S 34°52'W), João Pessoa, state of Paraíba, Brazil, 14–22.X.2003, S.C. Dias coll., deposited in IBSP 0 0 2. Paratypes, two females from the same locality and date, deposited in IBSP 0 0 3 and IBSP 0 0 4.
Etymology. The specific name honours the arachnologist Paula Frassinete Lins Duarte in recognition of her dedication to environmental conservation in northeastern Brazil, including her contribution to the establishment of the Mata do Buraquinho as a Forest Reserve.
Diagnosis. The male of Rowlandius linsduarteae can be distinguished from all other species of the genus by the combination of a pedipalp not elongated, without a ventral projection on trochanter ( Fig. 4 View FIGURES 1 – 6 ); the dorsoposterior process of opisthosomal segment XII small and subtriangular and the flagellum subquadrate, with dorsal projections located at the middle ( Fig. 1 View FIGURES 1 – 6 ) and with one ventral terminal spine ( Figs. 2–3 View FIGURES 1 – 6 ). Females resemble R. sul Cokendolpher & Reddell, 2000 by the presence of stalked lateral lobes and conical median lobes of spermathecae ( Cokendolpher & Reddell 2000: fig. 6). They can be differentiated by the absence of porous sculpturing on the spermathecal lobes and the irregular shape and convoluted inner space of terminal bulbs of lateral lobes in R. linsduarteae ( Fig. 6 View FIGURES 1 – 6 ).
Description. Male ( holotype). Anterior half of propeltidium, chelicerae, pedipalpi and leg I red brown. Rest of prosoma and remaining legs greenish brown. Venter of prosoma yellow. Opisthosoma dark brown, including flagellum. Propeltidium long, with two setae on anterior process and a median and a posterior pair of setae. Eye spot inconspicuous, elongate. Metapeltidium divided by a thin line. Anterior sternum with 12 setae. Opisthosomal tergites I–IX with a dorsal pair of setae each. Tergites X–XI narrow, with a lateral pair of setae. Tergite XII with a dorsal and two lateral pairs of setae. Segments XI-XII telescoped. Posterodorsal apophysis wide, subtriangular. Flagellum deeply excavated between dorsal projections, slightly elevated anteriorly ( Fig. 3 View FIGURES 1 – 6 ). Cheliceral basal segment with apical margin covered by long, white setae and with a serrated projection with one large apical tooth, a basal smaller one and several denticles in between. Distal segment straight and gently curved apically, with well developed serrula and guard tooth. Pedipalpal trochanter rounded, not projected anteriorly ( Fig. 4 View FIGURES 1 – 6 ). Leg tarsal segments proportion 2:1:2. Femur IV robust, anterior margin produced dorsally, approximately two times longer than high. Pedipalp measurements: trochanter 0.25/ femur 0.4/ patella 0.45/ tibia 0.35/ telotarsus 0.2/ total 1.65. Leg measurements: I trochanter 0.25/ femur 0.75/ patella 1.0/ tibia 0.7/ basitarsus 0.2/ telotarsus 0.4/ total 3.3; II 0.15/ 0.55/ 0.3/ 0.35/ 0.3/ 0.3/ 1.95; III 0.15/ 0.5/ 0.15/ 0.2/ 0.25/ 0.3/ 1.25; IV 0.2/0.8/ 0.4/ 0.55/ 0.5/ 0.35/ 2.8. Total length 2.9. Propeltidium 0.95 long, 0.45 wide. Prosoma 1.25 long. Opisthosoma 1.65 long (flagellum excluded).
Female ( paratype IBSP 003). Color, shape, chelicerae ( Fig. 5 View FIGURES 1 – 6 ) and spination as in male. Pedipalp not sexually dimorphic. Pedipalp measurements: trochanter 0.3/ femur 0.3/ patella 0.4/ tibia 0.35/ telotarsus 0.2/ total 1.55. Leg measurements: I trochanter 0.25/ femur 0.8/ patella 0.95/ tibia 0.6/ basitarsus 0.2/ telotarsus 0.35/ total 3.15; II 0.15/ 0.5/ 0.35/ 0.4/ 0.35/ 0.3/ 2.05; III 0.15/ 0.55/ 0.25/ 0.25/ 0.3/ 0.3/ 1.8; IV 0.25/ 0.8/ 0.35/ 0.5/ 0.4/ 0.3/ 2.6. Total length 3.35. Propeltidium 0.95 long, 0.5 wide. Prosoma 1.4 long. Opisthosoma 1.95 long (flagellum excluded). Flagellum with four segments. Internal genitalia without gonopod. Median lobes of spermathecae small and membranous. Lateral lobes with a long, curved stalk ( Fig. 6 View FIGURES 1 – 6 ).
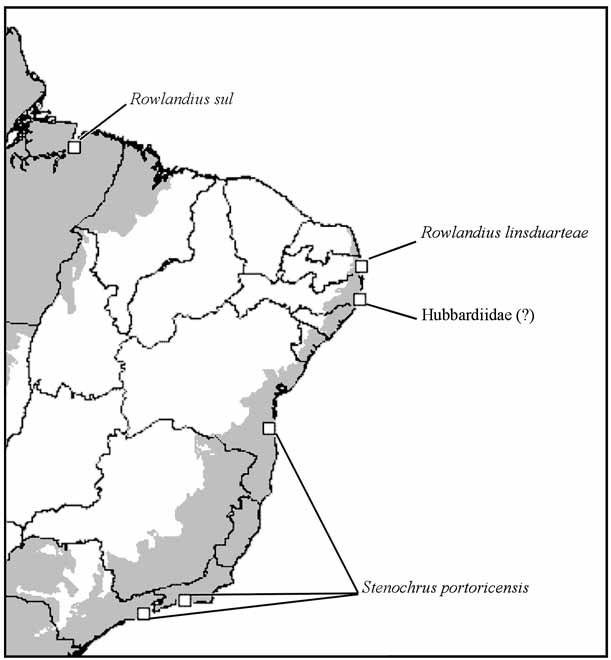
Distribution. Known only from type locality ( Fig. 9 View FIGURE 9 ).
Natural history. All the specimens examined were collected with pitfall traps in a secondary forest with dense undergrowth foliage.
Remarks. The genus Rowlandius Reddell & Cokendolpher, 2005 currently includes 50 species ( Harvey 2008), almost all of them restricted to the West Indies, mainly Cuba and Hispaniola ( Reddell & Cokendolpher 1995, Harvey 2003, Armas 2004). The only species previously known from South America is R. sul , described based on a female from eastern Brazilian Amazonia ( Cokendolpher & Reddell 2000, see Fig. 9 View FIGURE 9 ). This species is possibly closely related to R. linsduarteae , judging by the structure of the female genitalia. Both species share the absence of a gonopod, the lateral lobes of spermathecae with long and curved stalks and the median lobes small and conical. Compared to other species of the genus and of other genera of Hubbardiidae ( Rowland & Reddell 1978, 1979, 1981, Reddell & Cokendolpher 1995, Armas 2004), these characters are apparently derived and thus are potentially synapomorphic for R. linsduarteae and R. sul .
The biota of the northern portions of the Atlantic Forest is markedly different from the southern areas ( Silva et al. 2004, Santos et al. 2007) and has been considered as closer to the Amazonian biota, from which it was separated by semi-arid and savanna-like vegetation biomes ( Hueck 1972, see Fig. 9 View FIGURE 9 ). Using a Parsimony Analysis of Endemicity, Santos et al. (2007) indicate that the arboreal flora of forest enclaves of the northeastern Brazil is closely related to the Amazonian forest sites. These results suggest an ancestral northern connection between the two large Brazilian forest areas, followed by vicariance promoted by isolation by xeric vegetation. There are no similar studies for the fauna of northeastern Atlantic Forest, which have not been included in biogeographic analysis of South American lowland forests (e.g. Ron 2000, Pinto-da-Rocha et al. 2005). However, a phylogeographic analysis of didelphid marsupials also supports an ancestral connection between eastern Amazon and the northeastern Brazilian Atlantic Forest ( Patton & Costa 2003). Additional evidence comes from a recent revision of the Neotropical harvestmen family Stygnidae , in which Protimesius evelinae (H. Soares & B. Soares, 1978) is reported from Amazonian forest areas in the states of Pará and Maranhão and in an Atlantic Forest locality in the northeastern state of Paraíba (Pinto-da-Rocha 1997). This hypothesis is reinforced by the current study, although the evidence from the schizomid is based on the premise that R. linsduarteae and R. sul are sister-groups (as argued above), which could be tested in the future with a cladistic analysis of the genus. This proposition will also depend on an improvement of the knowledge of the schizomid fauna of Atlantic Forest, since more undescribed species can still be found.
As mentioned above, a schizomid specimen has been previously reported from a northeastern Atlantic Forest locality (see “ Hubbardiidae (?)” in Fig. 9 View FIGURE 9 ). This specimen was collected with a pitfall trap in the Parque Estadual de Dois Irmãos, Recife, state of Pernambuco ( 7o55’S 34o52’W) and deposited in the Museu Nacional do Rio de Janeiro, Rio de Janeiro, Brazil ( Peres et al. 2006). However, the specimen was severely damaged, preventing any further identification besides family-level (A. Giupponi, personal communication). This specimen could be a member of R. linsduarteae , considering the proximity between Recife and João Pessoa ( Fig. 9 View FIGURE 9 ) and the fact that both are part of the same center of endemism, composed of several forest enclaves in northeastern Brazil ( Tabarelli et al. 2006). New collections on Parque Estadual de Dois Irmãos will be necessary to clarify the identity of that schizomid specimen.

| IBSP |
Instituto Biologico de Sao Paulo |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |