
Cyphacolus Priesner
|
publication ID |
https://doi.org/ 10.5281/zenodo.276183 |
|
DOI |
https://doi.org/10.5281/zenodo.6210253 |
|
persistent identifier |
https://treatment.plazi.org/id/03D40C35-FFC0-FFD0-98DA-D09106A0A8F7 |
|
treatment provided by |
Plazi |
|
scientific name |
Cyphacolus Priesner |
| status |
|
Cyphacolus Priesner View in CoL
Cyphacolus Priesner, 1951:123 View in CoL . Original description. Type: Cyphacolus veniprivus Priesner View in CoL , by monotypy and original designation. Muesebeck & Walkley, 1956: 345 [citation of type species]; Kozlov 1971: 36 [keyed, citation of type species]; Masner 1976: 64 [tribal placement]; Johnson 1992: 366 [catalog of world species]; Austin & Field, 1997: 15 [revised tribal placement, host record]; Austin & Iqbal, 2005: 18 [diagnosis and key to species].
Zoobank registration: urn:lsid:zoobank.org:act:85B4E914-30E6-481C-8678-766A449E5B62
Diagnosis. Cyphacolus can be distinguished from all other genera of Platygastroidea by the following combination of characters: metasoma pedunculate in shape; first metasomal tergite with laterally compressed horn (elliptical in cross-section); fore wing contoured to convex surface of metasoma (elytriform), distal venation absent but with dark infuscate patch at fore wing margin; propodeum with pair of large, spine-like flanges that flank metasomal horn.
Description. Body mainly appearing smooth and shiny, often with dichromatic markings.
Head. In anterior view suboval or subtriangular, not elongate in buccal region; female antenna 7- segmented, clava large and unsegmented or with indistinct suture lines; male antenna short, 11-segmented but appearing to be 10–segmented as last two funicle segments are closely joined, distal funicular segments becoming progressively broader so that antenna is almost subclavate.
Mesosoma . Notauli usually present as distinct grooves reaching no more than about half way to anterior margin of mesoscutum, sometimes hidden by coarse longitudinal sculpture; mesoscutellum either flat and transverse, with posterior margin usually straight in middle part, or dorsally convex and semicircular or oval in shape; propodeum with pair of broad, elongate spines which are blunt or truncate apically; macropterous, never brachypterous, fore wing narrow basally with transverse fold about 1/3 distance from body, broad in apical half, remainder of fore wing surface sinuate, convex apically, molded to dorsal surface of metasoma; fore wing venation lacking except submarginal vein, pronounced infuscate patch at position of marginal (C+R) and stigmal veins (r-rs) and sometimes elsewhere. Vein r-rs absent.
Metasoma. Pedunculate in dorsal view, i.e. T1 square or rectangular (rarely more transverse) with parallel or slightly curved lateral margins, metasoma otherwise broad, oval, in lateral view dorsal surface slightly to strongly convex; T1 with large, laterally compressed horn-like process (i.e. ellipsoidal in cross-section); T3 slightly longer than T2, sometimes subequal in length with T2; ovipositor at least 1.5x length of metasoma, with shaft curled back on itself within rounded head of the horn on T1; gonoplacs elongate, approximately 0.75x length of metasoma.
Comments. The taxonomy and likely phylogenetic affinities of the genus have been discussed by Austin & Iqbal (2005). The genus is putatively monophyletic based on the unusual shape and venation of the fore wing, and the shape of the metasomal horn, the latter character otherwise found only in Odontacolus Kieffer. Cyphacolus + Odontacolus clearly form a monophyletic group within the Baeini , based on the metasomal horn, presence of large blunt spines on the propodeum, and the pedunculate metasoma. As discussed by Austin & Iqbal (2005), the shape of the horn is apparently linked to the functional mechanics of the ovipositor system, as the ovipositor is retracted within the metasoma and is curled around in the curved head of the horn so that it forms an elongated, bent U-shape ( Austin 1983). However, although Cyphacolus is putatively monophyletic, Odontacolus does not possess any obvious synapomorphies, and thus may be paraphyletic with respect to the former genus. We are currently revising the Old World species of Odontacolus and, as part of that study, we intend to examine this question by undertaking a morphological phylogenetic analysis including a broad sampling of members of both genera.
More problematic is the position of Cyphacolus and Odontacolus within the Baeini . This is an extremely diverse tribe of platygastroid wasps with generally uncertain relationships, whose members are presumed to be all endoparasitoids of spider eggs ( Austin 1985; Austin & Field 1997; Austin et al. 2005). More recently, one Cyphacolus Australian species was reared from an unknown spider’s egg sac, potentially confirming this trend for the Baeini clade.
The preliminary analysis of Iqbal & Austin (2000) employing 35 morphological characters, placed these two genera within a basal clade along with Hickmanella Austin and several undescribed species of Idris Foerster s. str. and Ceratobaeus Ashmead , the latter possessing a metasomal horn and being treated as a synonym of Idris by some authors (see Johnson 1992). However, the results of this analysis lacked stability because of the extremely high level of reductional synapomorphies (i.e. character losses) displayed by the Baeini , particularly the reduction of wing length and convergence of the highly fusiform body shape associated with cryptobiotic habits of some taxa, typified by Baeus Haliday (Austin et al. 2005; Stevens & Austin 2007). Interestingly, these reductional states do not occur in Cyphacolus and Odontacolus , and they show no apparent specializations for searching out host egg sacs in leaf-litter, with possibly one exception, i.e., the contoured to convex surface of metasoma-like fore wings of some Cyphacolus . This character is strongly reminiscent of the situation in the ambositrine diapriid genus Acanthobetyla Dodd , where the contoured fore wings of the female also lack venation and the wings are heavily sclerotised and deeply infuscate ( Naumann 1982). This arrangement presumably protects the surface of the fore wings when females are moving through litter, and the same may also apply to members of Cyphacolus , although at this stage we know very little about the biology of this genus.
Recently, Carey et al. (2006) examined the phylogeny of the Baeini using sequence data from the nuclear 28S rRNA and mitochondrial CO1 genes for an exemplar set of 21 ingroup taxa. Although preliminary in nature given the immense diversity of the tribe, the results of this study are interesting in that they show 1) the tribe is only monophyletic with the exclusion of the Australian genus Mirobaeoides Dodd , 2) that Baeus is the sister taxa to all remaining Baeini , 3) that Odontacolus is sister to a clade comprising Hickmanella Austin , Idris and Ceratobaeus , and 4) that neither of the latter two genera are monophyletic. Nonetheless, this study did not include any Cyphacolus species.
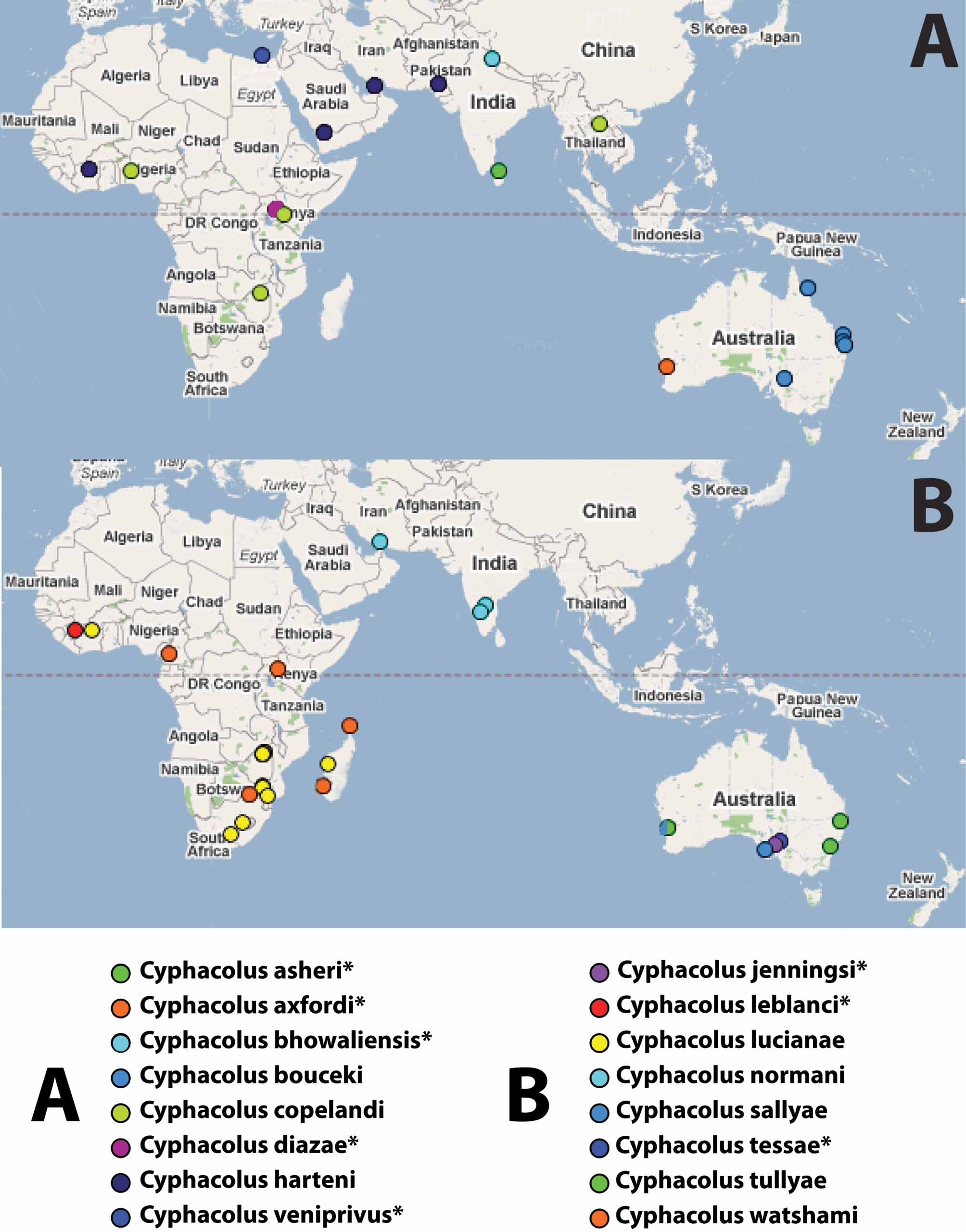
Link to distribution map. 16 Cyphacolus is found throughout the Old World tropics and subtropics, but extends into seasonally arid and semiarid regions outside of the tropics. It is found in South, West and East Africa ( South Africa, Zimbabwe, Cameroon, Guinea, Ivory Coast, Kenya), Madagascar, the Middle East ( Egypt, Yemen, United Arab Emirates), Pakistan, India, Sri Lanka, Thailand and Australia. It has so far not been recorded from south-east Asia south of latitude 16.5°N but very likely has a continuous distribution between India and northern Australia ( Fig. 2 View FIGURE 2 ).
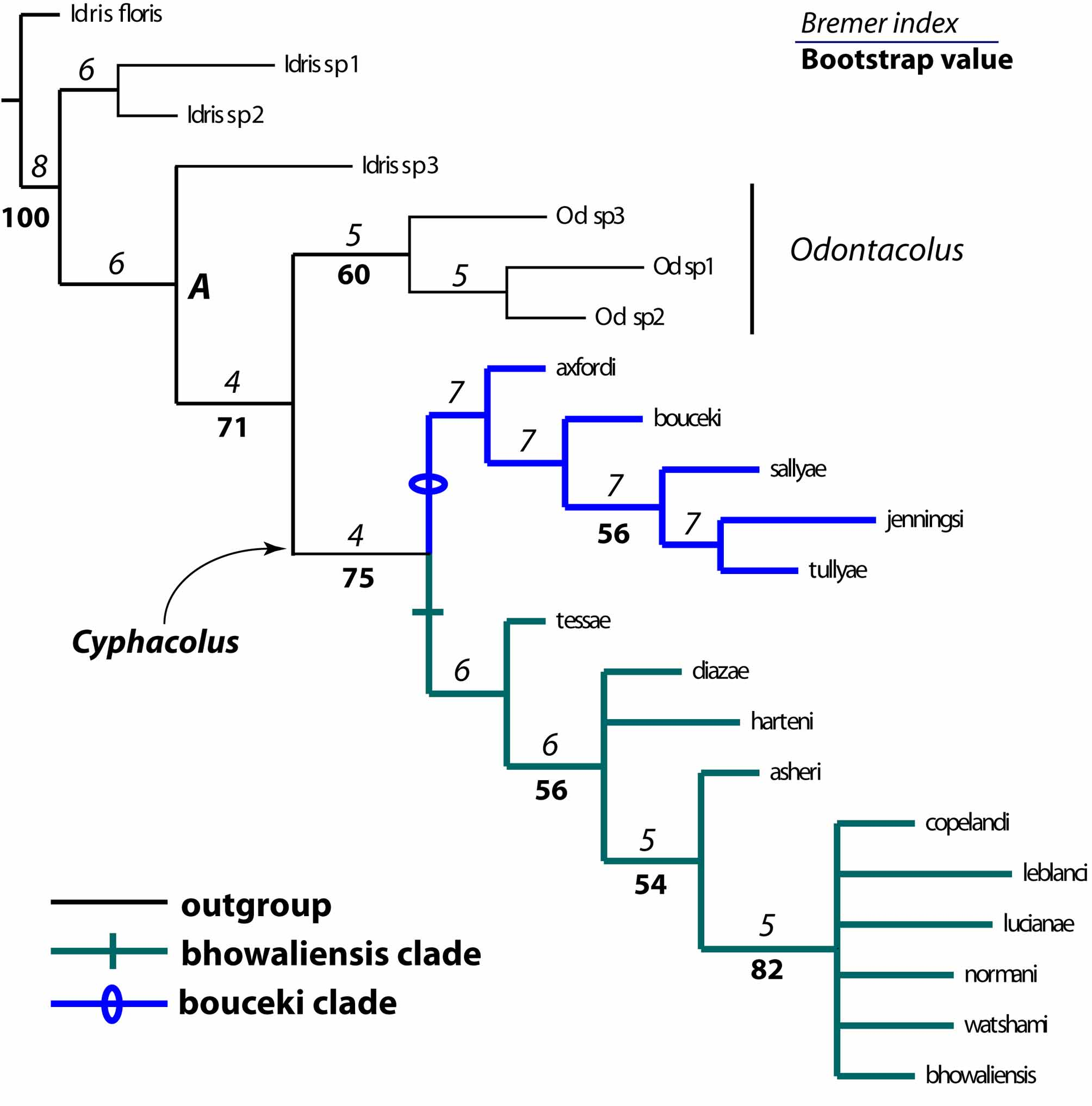
16. http://osuc.biosci.ohio-state.edu/HymOnline/map-large.html?id=468 Relationships among species. The phylogenetic analysis found eight trees of 141 steps from which a strict consensus tree was calculated (CI=0.530, RI=0.663, Fig. 3 View FIGURE 3 ). Cyphacolus was confirmed to be monophyletic (bootstrap = 75; Bremer = 4), as was Odontacolus (bootstrap = 60; Bremer = 5) which was also recovered as sister to Cyphacolus . Monophyly of Cyphacolus is supported by four character-states: lack of a facial keel, glabrous compound eyes, lack of a stigmal vein (r-rs) in the forewing, and the convexity of the fore wings. The main characters supporting the monophyly of Odontacolus are the longer mid-mandibular tooth when compared with the upper and lower teeth, the very setose compound eyes ( Cyphacolus compound eyes are glabrous), and the presence of a well-defined netrion. Although Odontacolus is resolved here as monophyletic we have included only three of an estimated 40 species for the genus, and further testing of the monophyly of the genus is needed with a broader set of Odontacolus species.
Within Cyphacolus two major clades are resolved which we attribute here to species groups. The bouceki species group is composed of C. axfordi , C. bouceki , C. sallyae , C. jenningsi , and C. tullyae . This clade is supported by the mainly smooth metasomal horn having some longitudinal costate sculpture on the posterior edge. This group contains only species from Australia, with C. tessae being the only taxon from the continent not included in this group.
The bhowaliensis species group includes C. tessae , C. hardeni , C. diazae , C. asheri , C. copelandi , C. lucianae , C. normani , C. watshami and C. bhowaliensis . This clade is supported by the presence of posterior lateral carinae on the metasomal horn, and by the absence of sculpture on the posterior area of the horn. With the exception of C. tessae (from Australia), all species comprising this group are found from Africa to India, with a single specimen of C. copelandi known from Thailand.
Within the bhowaliensis species group the relationships among several species are noteworthy. The species C. tessae , C. diazae and C. harteni do not have longitudinal costate sculpture on the mesoscutum, a character that is present for the remainder of species within the bhowalensis clade. Additionally, C. diazae and C. harteni can be separated from C. tessae by the occipital carina which is well separated from the orbital carina. Also contained within the bhowaliensis group is a tightly knit group of species comprising C. asheri and the six taxa in the unresolved clade at the apex of the tree ( Fig. 3 View FIGURE 3 ). These conspicuous species are united by having an unusual and pronounced globular mesoscutellum and the notauli absent. Members of the unresolved clade (comprising C. copelandi , C. lucianae , C. normani , C. watshami and C. bhowaliensis ) are further distinguished by a range of character states including antennal scrobes and malar space with confused sinuate transverse costate sculpture throughout, and the propodeal spines with lobate areas apically and being wider apically than at their base. This group of species has the metasoma very strongly convex and normally polished dorsally, and the anterior margin of the fore wing sinuate just basal of the mid-point. Cyphacolus asheri , according to the present analysis, potentially belongs to this group but currently is known only from a single male, and there is the possibility that the female will show a different morphology based on the sexually dimorphic nature of this genus. The exclusion of C. asheri from the analysis hardly affects the relationships depicted in Figure 3 View FIGURE 3 , the only significant change being that C. harteni is basal to C. diazae .
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Cyphacolus Priesner
| Valerio, Alejandro A., Masner, Lubomir & Austin, Andrew D. 2010 |
Cyphacolus
| Austin 2005: 18 |
| Austin 1997: 15 |
| Johnson 1992: 366 |
| Kozlov 1971: 36 |
| Muesebeck 1956: 345 |
| Priesner 1951: 123 |