
Lamprometra palmata ( Müller, 1841 ), Muller, 1841
|
publication ID |
https://doi.org/10.5281/zenodo.184295 |
|
DOI |
https://doi.org/10.5281/zenodo.6227275 |
|
persistent identifier |
https://treatment.plazi.org/id/03D4870E-FFF1-770C-FF6C-FBF5BAF81E59 |
|
treatment provided by |
Plazi |
|
scientific name |
Lamprometra palmata ( Müller, 1841 ) |
| status |
|
Lamprometra palmata ( Müller, 1841) View in CoL
Alecto palmata Müller, 1841:185 View in CoL .
Comatula ( Alecto) palmata: Müller, 1847:261 .—Lütken, 1874:190 (in Carpenter, 1879).
Antedon protectus Lütken, 1874:190 (part) ( nomen nudum, in Carpenter, 1879, p. 18–19).—AH Clark, 1913b:31.
Antedon palmata: Carpenter, 1879:23 (part), 29 (part), 45; 1882:733; 1888:35 (part), 44 (part), 54 (part), 55 (part), 208 (part), 213 (part), 366 (part), 379.—AH Clark, 1912c:385.
Antedon protecta Carpenter, 1881:192 ; 1888:53, 54, 91, 225, 234, 237, 366, 379.— Hartlaub, 1890:170, 180.
Antedon gyges Bell, 1884:160 –161, pl. 12, fig. b.— Carpenter, 1888:49 –50, 55, 224–225, 366, 379.—AH Clark, 1911a:714, 716.—HL Clark, 1921:6.
Antedon conjungens Carpenter, 1888:233 , pl. 1, fig. 1; 1889:305, 311, pl. 27.— Bell, 1888:389.—AH Clark, 1941:482 – 483.
Antedon klunzingeri Hartlaub, 1890:175 ; 1891:46, pl. 2, figs. 22, 25; 1912:152.—AH Clark, 1909c:4.
Antedon protectus : AH Clark, 1908b:489.
Lamprometra protectus View in CoL : AH Clark, 1913b:31, 179; 1918:100, pl. 8, pl. 27, figs. 99, 100.— Gislén, 1922:75.
Lamprometra brachypecha View in CoL HL Clark, 1915b:104; 1921:8, 22: 192 f., pl. 2, fig. 1, pl. 22, figs. 1, 2.—AH Clark, 1941:489 –90.
Lamprometra protecta View in CoL : HL Clark, 1921:23, 192.—AH Clark, 1929:641.
Lamprometra gyges View in CoL : AH Clark, 1913a:144.—HL Clark, 1921:23, pl. 1, fig. 4, pl. 21, figs. 4, 5, pl. 22, fig. 3.— Gislén, 1934:25.
Lamprometra klunzingeri View in CoL AH Clark, 1913a:144.
Lamprometra palmata View in CoL : AH Clark, 1929:641.— Gislén, 1936:2, 4–5, 12, 14.— HL Clark, 1946:47.—AM Clark, 1975:402.—AM Clark & Rowe, 1971:23, fig. 8b.— Meyer & Macurda, 1980: 84 –85, figs. 5c, 5d, 7a.— Zmarzly, 1985:352.— Chen et al., 1988:78, fig. 21.— Stevens, 1989:4 –20, 4–21, pl. 7a, 4–28, 6–14, figs. 6.10–6.13, 6–20, fig. 6.8.— Messing, 1994:238 –239; 1998:189, 190.— Kogo, 1998:65 –67, fig. 53.
Stephanometra oxyacantha View in CoL : AH Clark, 1941:421 (one misidentified specimen).
Lamprometra palmata palmata View in CoL : AH Clark, 1941:474 –517, pl. 53, figs. 243–246, pl. 54, figs. 248–252, pl. 55, figs., 257.—AM Clark, 1972:104 –105, fig. 10 (a–e).
Lamprometra palmata gyges View in CoL : AH Clark, 1941:517 –526, figs. 253–255. Lamprometra parmata parmata Utinomi & Kogo, 1965:274 –276, fig. 7. Stephanometra spicata: Kogo, 1998:61 View in CoL –63, fig. 49.
Holotype.— Alecto palmata Müller, 1841 . AH Clark recorded the specimen, collected by D.F. Eschricht in India, as in the Berlin Museum (1908b:452) (=Museum für Naturkunde der Humboldt-Universität zu Berlin) or the Anatomical Museum in Berlin (1941:511) (either Das Berliner Medizinhistorisches Museum der Charité, or Charité - Universitätsmedizin Berlin). It cannot be found in the collections of any of these institutions and is apparently lost.
Material examined. RED SEA: USNM 35224 (1 spec.), Misharif I., Khor Dongola, no additional data; USNM 35769 (1), Um el Jerman, Gulf of Suez, R Hartmeyer, coll., no additional data. THAILAND: USNM E11630 (1), Andaman Sea, W of Phuket, 0 7°34'N, 98°00'E, 77 m, 21 Mar 1963. SINGAPORE: USNM E34853 View Materials (1), DL Meyer, coll., no additional data. INDONESIA: USNM E389 (1), Siboga 279, Ramah-Kuda Bay, 36 m, 11–13 Dec 1899; USNM E405 (1), Siboga 209, Kabaena I., Greater Sunda Is., 22 m, 23 Sep 1899; USNM E35050 (1), S end Banda I., Banda Sea, 2–3 m, 31 Jan 1975, DL Meyer, coll.; USNM E35370 View Materials (1), Rumphius II SEL-2, NW end Seleman Bay, Ceram Sea, 0 2°53'50"S, 129°05'15"E, 20 Jan 1975; USNM E36145 View Materials (2), Weda Bay, Halmahera I., 0 0°50'00"N, 127°34'00"E, 1.5–6 m, 14 July 1979, G Hendler, coll. MALAYSIAN BORNEO: NSUOC 360, 361, 363, FLMNH 6833 (4), Dive Center, Mabul I., 0 4°15'N, 118°38'E, 12 m, 24 Apr 1997, CG Messing coll.; FLMNH 6835, 6836, 6866, NSUOC 354, 357, 362 (6), E side Mabul I., 0 4°15'N, 118°38'E, 10–21 m, 22–26 Apr 1997, CG Messing, coll. PHILIPPINES: NSUOC 347 (1), W end S Tubbataha Reef, 0 9°49'N, 119°52'E, 21 m max., Apr 1995, CG Messing, coll; NSUOC 345 (1), Fondeado Reef, 24 Apr 1995, CG Messing, coll.; USNM 35252 (2), Albatross sta.?, no additional data; NHM 1888.11.9.76 (2), Challenger, Cebu Reef, no additional data. PALAU: CRRF 1151L (1), seagrass bed E entrance to Malakal Harbor, 0 7°17.90'N, 134°28.34'E, 1 m, 28 Jun 1993. CHUUK ATOLL: NSUOC 368 (1), Wreck of Fujikawa Maru, between Dublon and Uman Is., 0 7°20'N, 151°53'E, 9.1–12.1 m, 14 June 1993, CG Messing, coll. MARSHALL ISLANDS: USNM 35248 (1), Ebon I., Ralik Chain, 14 Apr 1877, BG Snow, coll., no additional data. PAPUA NEW GUINEA: NSUOC 358 (1), Barrier I. outside Magic Pass, Madang, 6– 8 m, 9 July 1991, L Harris, coll.; NSUOC 353 (1), Barracuda Rock, off Pig I., Madang, 0 5°10'20"N, 145°51'53"E, 6 m, 8 July 1991, CG Messing, coll.; FLMNH 6844 (1), outside Pig I., 0 5°10'20"N, 145°51'53"E, 11 m, 16 July 1991, CG Messing, coll.; NSUOC 344, 346 (2), Jais Aben Reef, N side Nagada Harbor, 0 5°09'29"S, 145°49'21''E, 3–4 m, 2 June 1992, CG Messing, coll.; NSUOC 352 (1), Barrier I. outside Wongat I., 0 5°08'09"S, 145°50'51"E, 3 m, 11 July 1991, CG Messing, coll.; FLMNH 6828 (1), Banana Rock, off Nagada Harbor, 0 5°9'29"S, 145°49'21"E, 6 m, 6 July 1991, L Harris, coll.; IRSNB / CRI 410, 411 (2), Hansa Bay, Laing I., sta. 13, 5 m, 24 June 1989, MC Lahaye, coll.; IRSNB / CRI 412 (1), Hansa Bay, Epave (Sisimangun), 6 m, 7 July 1989, MC Lahaye, coll; IRSNB / CRI 413 (1), Platier (NE), 20 m, 16 July 1989, MC Lahaye, coll.; IRSNB / CRI 414, 415 (2), Hansa Bay, Laing I., (Pointe 0), 25 m, 22 July 1989, MC Lahaye, coll.; IRSNB / CRI 416–418 (3), Platier, Hansa Bay, 20 m, 23 July 1989, MC Lahaye, coll.; IRSNB / CRI 419 (1), Mandy Passage (Near Hansa Bay), 41 m, 24 July 1989, MC Lahaye, coll. AUSTRALIA: USNM E34827 View Materials (3), Lizard I., 1975, DL Meyer, coll., no additional data; USNM E34738 View Materials (1), Heron I., DL Meyer, coll., no additional data; USNM 35261 (1), Bowen, Port Denison, Queensland, no additional data. FIJI: USNM E34799 View Materials (1), Makuluva I., 18°11'36"S, 178°31'16"E, 27 Apr 1975, DL Meyer & B Carlson, coll.; NHM 1888.11.9.77 (1), 1888.11.9.78 (3), Challenger 174, Kandavu, 19°06'S, 178°18'E, 411 m, 3 Aug 1874.
Diagnosis. As for genus.
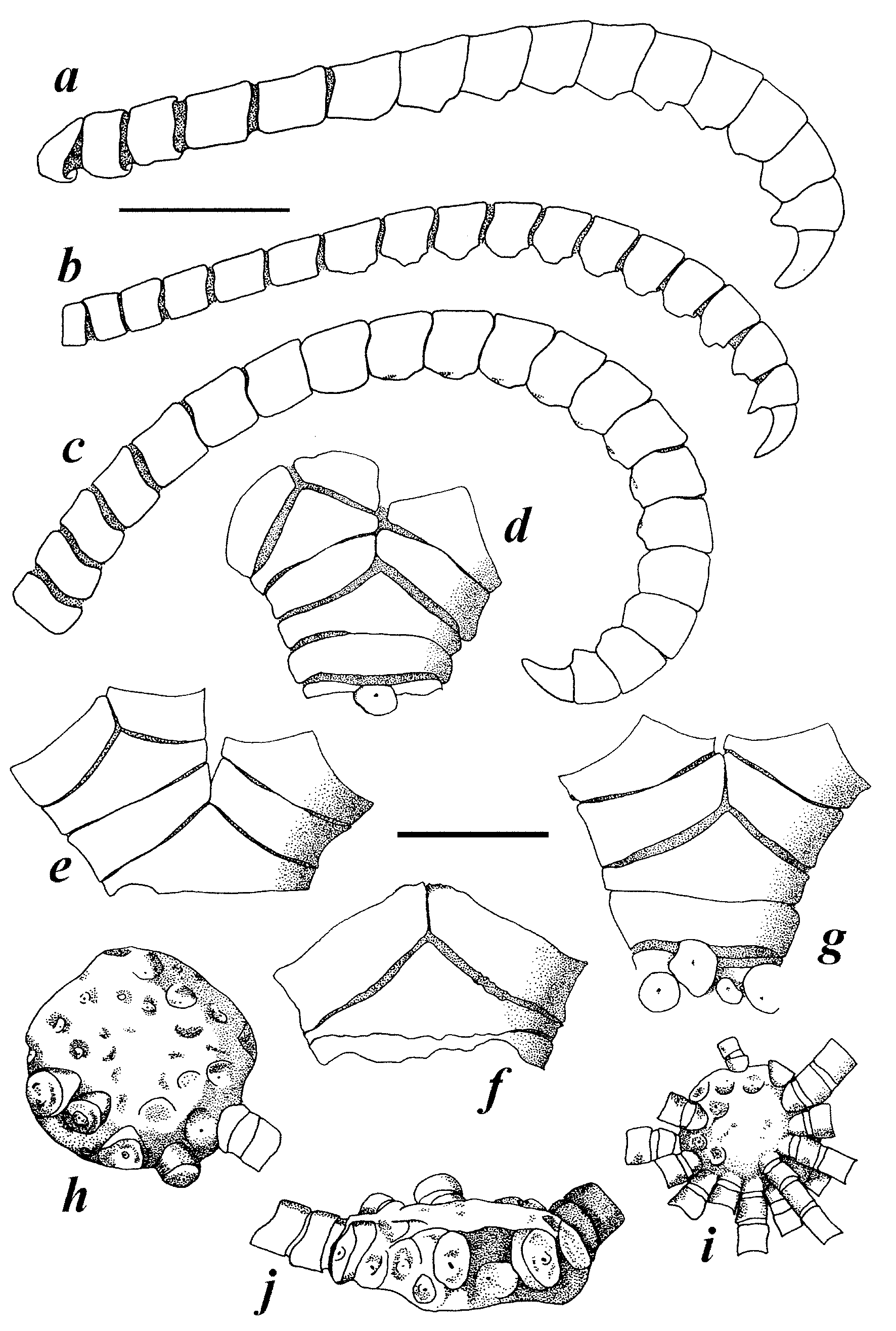
Description. Centrodorsal thin, discoidal ( Figs. 16 View FIGURE 16 h–j), 1.6–6.4 mm across, 0.6–1.9 mm high; DH 2.6– 5.7. Cirri in 1–2 or sometimes 3 alternating marginal rows. Polar area flat, bare, rarely convex, 0.9–4.6 mm across; DP 1.2–2.3. Cirri XVI–XLIV, 16–32, 9.0–21.0 mm long; c1–3 short; c4–6 slightly longer than broad; subsequent cirrals slightly broader than long; c6-8 longest, LW 0.9–1.6; c7–18 and following developing sharp aboral carination, becoming a low, swollen, triangular spine distally, sometimes worn into a low carination. Penultimate segment shorter than curved terminal claw and with a sharp spine ( Figs. 16 View FIGURE 16 a–c).
Basals not visible externally; radials sometimes visible at interradial angles. Brachitaxis ossicles ranging from strongly flattened against each other to separated ( Figs. 16 View FIGURE 16 d–g). Ibr1 commonly with diverging outer margins although sometimes with converging or parallel margins, usually united proximally, rarely free laterally; L to 1.1 mm, W to 3.7 mm; WL 2.1–5.8. Ibr2 with diverging lateral margins; L to 2.0 mm, W to 4.1 mm; WL 1.6–5.6. Lateral adambulacral margins of brachitaxis ossicles flattened, apposed, and weakly thickened, sometimes crenulated and interlocking, or smoothly rounded, neither apposed nor laterally thickened, with weak lateral adambulacral expansions. IIIBr2 developed externally only or externally and internally; IVBr2 either external or internal, if present. Synarthrial tubercles weak to well developed.
Arms 12–44; R 40–110 mm; rays usually with 2–4 or 6 arms each, less often with 5, 7, 8 or 9. Br1–br6 or br7 oblong; subsequent brachials cuneate. Br10 to 1.8 mm wide, 0.9 mm long, WL 1.2–3.2. Syzygies at br3+4, from br14+15 to br16+17, less often br9+10 and br17+18, and rarely at br12+13, br19+20 or br20+21. Distal syzygial intervals 7–11, less often 6 or 12.
Pinnules on outer arms generally longer than those on inner arms. P1-P3 composed chiefly of pinnulars barely longer than broad, and tapering gradually to a slender tip; terminal pinnular with numerous minute spines (Figs. 1e–h, 17). Basal three pinnulars oblong, laterally compressed, typically bearing a moderate to strong aboral carination (Fig. 17d) that may be completely reduced, especially on P2. Following pinnulars with LW 1.0–1.5. P1 3.0– 11.2 mm long, of 10–39 pinnulars, slender and shorter than P2, often slenderer and shorter than P3. P2 the longest, thickest pinnule, 4.3–23.0 mm long, of 12–41 pinnulars; pinnular 6 with LW 0.9–1.5, rarely greater. P3 2.1–9.1 mm, of 9–22 pinnulars, usually resembling P1 though sometimes slightly slenderer or thicker, sometimes small weak and flexible with a well-developed ambulacral groove as in the following pinnules. P4 1.7–5.9 mm, of 9–17 pinnulars. Following pinnules small, weak and flexible. P5 1.3–4.7 mm. Following pinnules gradually increasing in length. P distal 7.2 mm, of 18 pinnulars; the first pinnular broader than long, the second as long as broad; distal pinnulars barely longer than broad; the terminal segment elongated and covered with small spines.
Color patterns. Concentric banding on the arms formed by alternating colors in bands of 4–15 brachials each in various shades of white, cream or tan that alternate with dark brown, red-brown, rusty orange, pink or green. One specimen was banded purple and rusty orange. Specimens with rusty orange or green coloration were frequently observed with white brachitaxes blotched, respectively, with rusty orange or green. Additional patterns (e.g., AH Clark 1921b; Zmarzly 1985; Stevens 1989) may be based on incorrectly identified specimens.
Stevens (1989) noted that proximal arm colors were inconspicuous while distal portions tended to be “bright and striking” and suggested that this pattern provided camouflage when the arms curled in. Most of Meyer & Macurda’s (1980) specimens had a characteristic banded green and white chevron pattern. HL Clark (1915b:104) referred specimens with this color pattern to L. brachypecha . AH Clark (1921b) ascribed it to Lamprometra protectus , which he thought was the same as L. palmata .
Distribution. On reefs from the Red Sea and Mauritius in the west to Tonga in the east, and from tropical Australia to southern Japan (AH Clark 1941; Utinomi & Kogo 1965; Kogo 1998). Previously published records may be based on incorrectly identified species. A single fragmentary specimen from Oahu, Hawai’i, was ascribed to L. palmata (AH Clark 1941), but its presence in the Hawaiian chain has not been confirmed. Depth range: ~ 1 to 35 m. Two deeper records are likely errors: USNM E11630 (R/V Anton Bruun sta. 18A), 77 m; NHM 1888.11.9.78 (HMS Challenger), 411 m (4 spec.) (AH Clark 1941).
Ecology.—Cryptic on reefs during the day under coral rubble and slabs or within the reef infrastructure. At night, perched on prominent coral heads, branching corals, large rubble, coral rock projections or large sponges, with arms arranged in a biplanar arcuate fan, funnel or shallow bowl. At Palau, they emerge within an hour before dusk and crawl to the tops of coral perches within ~15 min ( Meyer & Macurda, 1980). Also at Palau, L. palmata is common among branching corals in a macroalgae and seagrass bed in 1 m (Ngadarak Bank, S side of Ngell Channel, 07º 17.90’N, 134º 28.34’E.). The species also forms an irregular radial fan in the open during the day in strong flow, but under reduced light conditions inside a tunnel to a marine lake ( Messing 2007). Stevens (1989) also reported that it forms partly exposed arm fans during the day in> 12 m at Heron and Wistari Reefs, southern Great Barrier Reef, Australia.
FIGURE 17. Lamprometra palmata , pinnules. a. NSUOC 345, P1-P3. b. NSUOC 360, P1-P3. c. NSUOC 368, P1-P3. d. USNM 1094083, proximal pinnulars of P2. e. NSUOC 346, L. palmata form brachypecha , P1-P4. Scales: upper (a–c), right (e) all 2 mm; middle (d) 0.5 mm.
Remarks. AH Clark (1941) distinguished Lamprometra from other Mariametridae by a P2 usually stouter and much longer than P3 and tapering to a slender delicate tip. The revised diagnosis above includes features noted by previous authors but not included as generic characteristics: centrodorsal aboral pole flat, broad and without cirrus sockets; P2 composed of pinnulars that are not especially elongated, and brachitaxes ossicles occasionally with crenulated exterior lateral margins. Diagnostic characters not previously noted include normally developed oral pinnular facets (Fig. 2e, f), terminal pinnulars of oral pinnules bearing spines (Fig. 1h), and a reduced ambulacral groove (as in Stephanometra ) on P1, P2 and sometimes P3.
Within the genus, AH Clark (1941) distinguished two species: L. klunzingeri from the Red Sea south to Oman and possibly Tanzania with slender proximal pinnules and basal pinnulars bearing a slight carination or none at all, and L. palmata from Baluchistan (now part of Pakistan) to the Pacific typically with P2 (and sometimes P3) markedly stouter than the other oral pinnules. However, he noted that L. klunzingeri might be just “the western form” (p. 473) of L. palmata . Within L. palmata , he formally recognized two morphs that he called both varieties and forms, though he treated them nomenclatorily as subspecific trinomens: L. p. palmata with P2 stouter than the other oral pinnules, and L. p. gyges ( Bell, 1884) with all proximal pinnules slen- der as in L. klunzingeri but with basal pinnulars strongly carinate. He maintained gyges as a distinct variety because of its “more or less distinctive geographic range,” though he acknowledged that it broadly overlapped that of L. p. palmata , and that the two intergraded completely and were often found together. He also provided a key to an additional 14 named forms of L. palmata previously considered separate species, noting that some were perhaps “worthy of recognition” as well-marked stable varieties, while most were “individual variants or growth or developmental stages” (p. 505). He did not identify which were which. As another source of confusion, this key distinguishes the gyges and palmata forms as having 30 versus not over 25 cirrals, although his description of L. p. gyges that follows refers to 20–30 (usually 20–25) cirrals. One specimen in the current study possesses more than 30 cirrals.
Although AM Clark & Rowe (1971) followed AH Clark’s (1941) taxonomy, they noted specimens from the Red Sea with strongly carinate oral pinnule bases characteristic of L. p. gyges and others from Australia with hardly a trace of a keel—characteristic of L. klunzingeri . As a result, they suggested that both klunzingeri and gyges might be better considered as subspecies of L. palmata . Specimens in the current study display similar discrepancies. Five of 52 measured specimens have P3 longer than P1 as in L. klunzingeri , but the basal oral pinnulars are carinate in all five, and none is from the Red Sea or western Indian Ocean. USNM E35769 View Materials from the Red Sea, identified by AH Clark as L. klunzingeri , has carinate basal pinnulars and P1 ( 10.7 mm) longer than P3 ( 6.8 mm), which contradict his description of the species. USNM E34717 View Materials from Malaysia, labeled L. klunzingeri , exhibits basal carination, P3 15 mm long with 30 pinnulars, and P2 10 mm with 26 pinnulars. We regard this specimen as Dichrometra sp.
Although specimens with separated versus strongly flattened and apposed adambulacral margins of brachitaxes exhibit strongly different overall appearances, a complete continuum between extremes exists. Bivariate plots of several characters (centrodorsal diameter and height, ray length, number of cirri and Ibr2 width) that increase with growth reveal no distinction between specimens with different margins except that those with apposed flattened margins reach a larger size.
Basal carination of oral pinnulars, form of brachitaxial adambulacral margins, and numbers of cirrals all vary substantially in Lamprometra . Specimens often could not be satisfactorily assigned to a species or subspecies based on these diagnostic features. We therefore treat L. klunzingeri and L. p. gyges as infrasubspecific variants of L. palmata , confirming AM Clark’s (1975) doubts about the worth of recognizing either specific or subspecific distinctions.
HL Clark (1915) described Lamprometra brachypecha as new based on its small size, shorter arms, fewer cirri, completely smooth oral pinnules and distinctive color. The holotype was bright green, variegated with brown and white, with a broad band of white on the arms, and yellow-tipped distal pinnules. He observed that the undisturbed animal, with arms covering its oral surface, resembled a tuft of green seaweed. When touched, the arms extended in a flat fan and a broad white band flashed into view. “The immediate effect was obliterative and one’s first thought was that the animal had vanished” (p. 104). Although AH Clark (1941) distinguished brachypecha by its short arms composed of approximately 100 brachials, he treated it as a form of L. palmata .
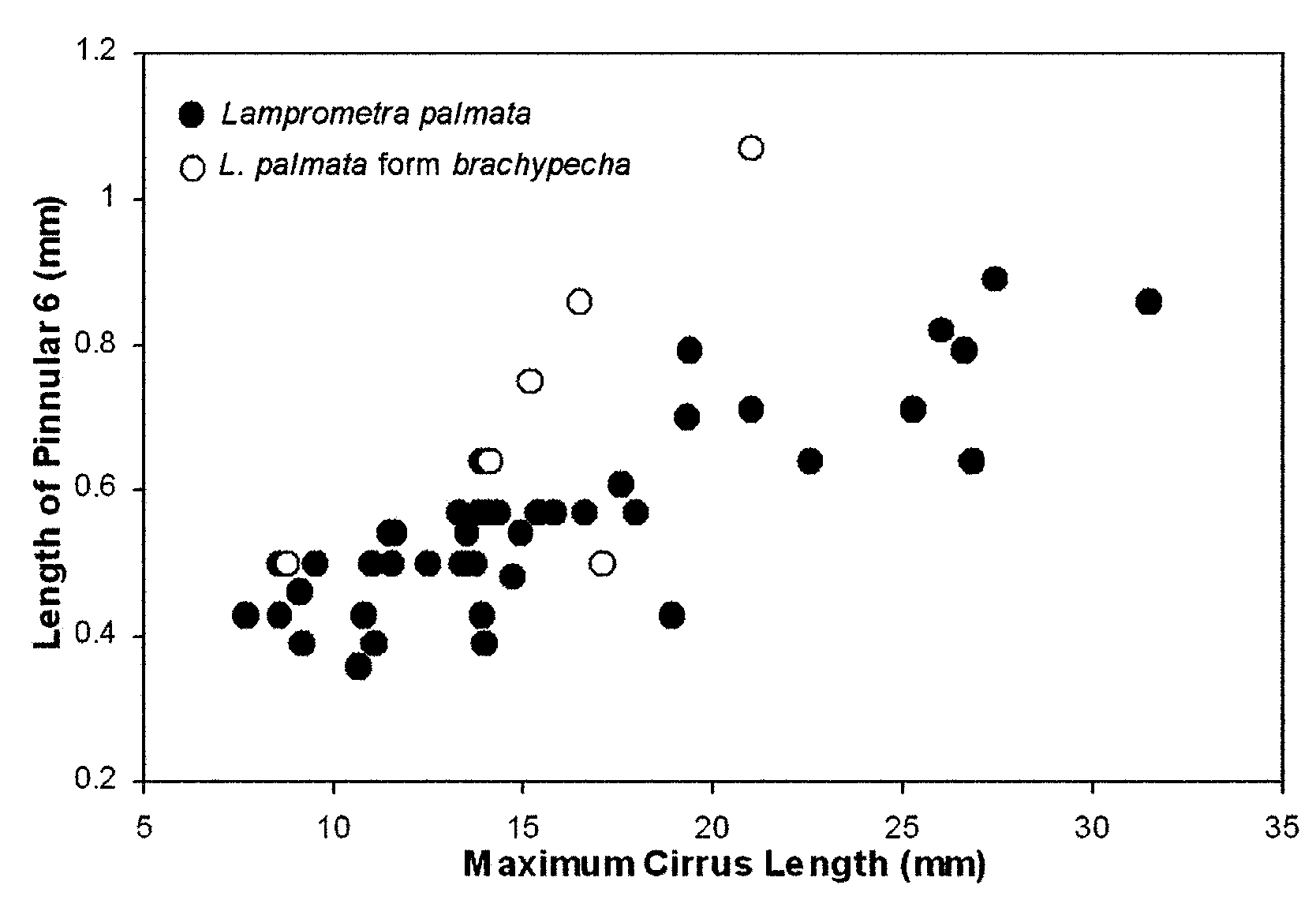
We originally identified six specimens in this study as L. palmata form brachypecha and singled them out as perhaps representing a distinct taxon based on their color and extremely thick, enlarged P2 (Fig. 17e). The centrodorsal and brachitaxes ossicles were white or cream mottled with green spots, and the arms banded dark green distally. P2 was both longer and stouter than in other specimens, up to 23.0 mm in length with 41 pinnulars and with a mean basal width of 0.70 mm (maximum 1.14 mm) relative to the mean of 0.45 mm in typical L. palmata . In bivariate plots of characters that vary with growth, L. p. form brachypecha falls well within the L. palmata character space (not shown). However, in plots of the length of P2 pinnular 6 against maximum cirrus length and Ibr2 width ( Fig. 18 View FIGURE 18 ), larger specimens of L. p. form brachypecha fall outside the L. palmata character space. However, one of us (CGM) subsequently collected or observed additional specimens at Palau with P2 typical of L. p. form brachypecha ( Messing 2007) . Of these, four had the typical green and white color pattern, but two differed: 1) ray bases white with rusty orange blotches; arms rusty orange with three pairs of narrow white bands, and arm tips brown and white; 2) ray bases white with purplish-gray blotches; mid-arms brown with 2–3 pairs of narrow white bands followed by successive sections of white, purplish-gray and white. Such variations reduce the likelihood that this represents a distinct taxon. However, a definitive determination requires additional specimens with this distinctive pinnule morphology supplemented by molecular sequence data.
The length of P2 relative to P3 is the only published character distinguishing Lamprometra from the mariametrid genera Liparometra and Dichrometra : P2>P 3 in Lamprometra ; P2<P 3 in Dichrometra , and P2=P 3 in Liparometra (AH Clark 1941). An examination of 28 specimens (from Chuuk Atoll, Australia, Malaysia, Philippines and Papua New Guinea) ostensibly belonging to the latter two genera have so far revealed no other distinguishing features. Lateral adambulacral margins, pinnular shape, cirrus form and centrodorsal shape are similar in all three. In addition, intermediates exist. In USNM E34827 View Materials (Heron I., Queensland, Australia) and FLMNH 6813 (Chuuk), P2 is longer and thicker than P3 on some exterior arms as in Lamprometra and equal to P3 on others as in Liparometra . In FLMNH 6850 ( Papua New Guinea) and IRSNB/CRI 420, P2 is equal to P 3 in length and thickness on some arms as in Liparometra and smaller on others as in Dichrometra . No regeneration was apparent in any of these. Both Gislén (1922) and HL Clark (1938) questioned the distinction among the three, and AH Clark (1941), though maintaining them as separate genera, acknowledged that some specimens could not be assigned to genus based on oral pinnule form. In Figure 19, two bivariate plots illustrate how representative specimens of the three genera plus intermediates occupy strongly overlapping character spaces, although both Liparometra and Dichrometra specimens reach larger sizes than Lamprometra palmata . We have not identified specimens of the former two genera to species, as substantial overlap exists among most of them. AH Clark (1941) acknowledged that two of his three species of Liparometra and at least three of the seven species of Dichrometra might represent single species. Similarly, we have not formally synonymized Liparometra and Dichrometra under Lamprometra pending a more detailed examination of component species.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Lamprometra palmata ( Müller, 1841 )
| Rankin, Dana L. & Messing, Charles G. 2008 |
Stephanometra oxyacantha
| Clark 1941: 421 |
Lamprometra palmata palmata
| Clark 1972: 104 |
| Clark 1941: 474 |
Lamprometra palmata gyges
| Kogo 1998: 61 |
| Utinomi 1965: 274 |
| Clark 1941: 517 |
Lamprometra protecta
| Clark 1929: 641 |
Lamprometra palmata
| Kogo 1998: 65 |
| Messing 1994: 238 |
| Stevens 1989: 4 |
| Chen 1988: 78 |
| Zmarzly 1985: 352 |
| Meyer 1980: 84 |
| Clark 1975: 402 |
| Clark 1971: 23 |
| Clark 1946: 47 |
| Gislen 1936: 2 |
| Clark 1929: 641 |
Lamprometra brachypecha
| Clark 1941: 489 |
| Clark 1915: 104 |
Antedon protectus Lütken, 1874:190
| Clark 1913: 31 |
Lamprometra protectus
| Gislen 1922: 75 |
| Clark 1913: 31 |
Lamprometra gyges
| Gislen 1934: 25 |
| Clark 1913: 144 |
Lamprometra klunzingeri
| Clark 1913: 144 |
Antedon protectus
| Clark 1908: 489 |
Antedon klunzingeri
| Clark 1909: 4 |
| Hartlaub 1890: 175 |
Antedon conjungens
| Clark 1941: 482 |
| Carpenter 1888: 233 |
| Bell 1888: 389 |
Antedon gyges
| Clark 1911: 714 |
| Carpenter 1888: 49 |
| Bell 1884: 160 |
Antedon protecta
| Hartlaub 1890: 170 |
| Carpenter 1881: 192 |
Antedon palmata :
| Clark 1912: 385 |
| Carpenter 1879: 23 |
Comatula ( Alecto ) palmata : Müller, 1847 :261
| Muller 1847: 261 |
Alecto palmata Müller, 1841 :185
| Muller 1841: 185 |