
Cyclaxyridae, Gimmel & Leschen & Ślipiński & Csiro, 2009
|
publication ID |
https://doi.org/ 10.5281/zenodo.5319990 |
|
persistent identifier |
https://treatment.plazi.org/id/03D48786-FFFE-EA2C-2992-9A728E97064E |
|
treatment provided by |
Felipe |
|
scientific name |
Cyclaxyridae |
| status |
fam. nov. |
Cyclaxyridae View in CoL View at ENA , fam. nov.
Cyclaxyrinae Klimaszewski & Watt 1997: 49, as subfamily of Phalacridae Leach, 1815 View in CoL . Nomen nudum. Cyclaxyridae Lawrence et al. 1999a View in CoL . Unavailable name. Cyclaxyridae Lawrence & Leschen 2003: 910 View in CoL . Nomen nudum. Cyclaxyridae Leschen et al. 2005: 63 View in CoL , 67. Nomen nudum.
Type genus. Cyclaxyra Broun, 1893 .
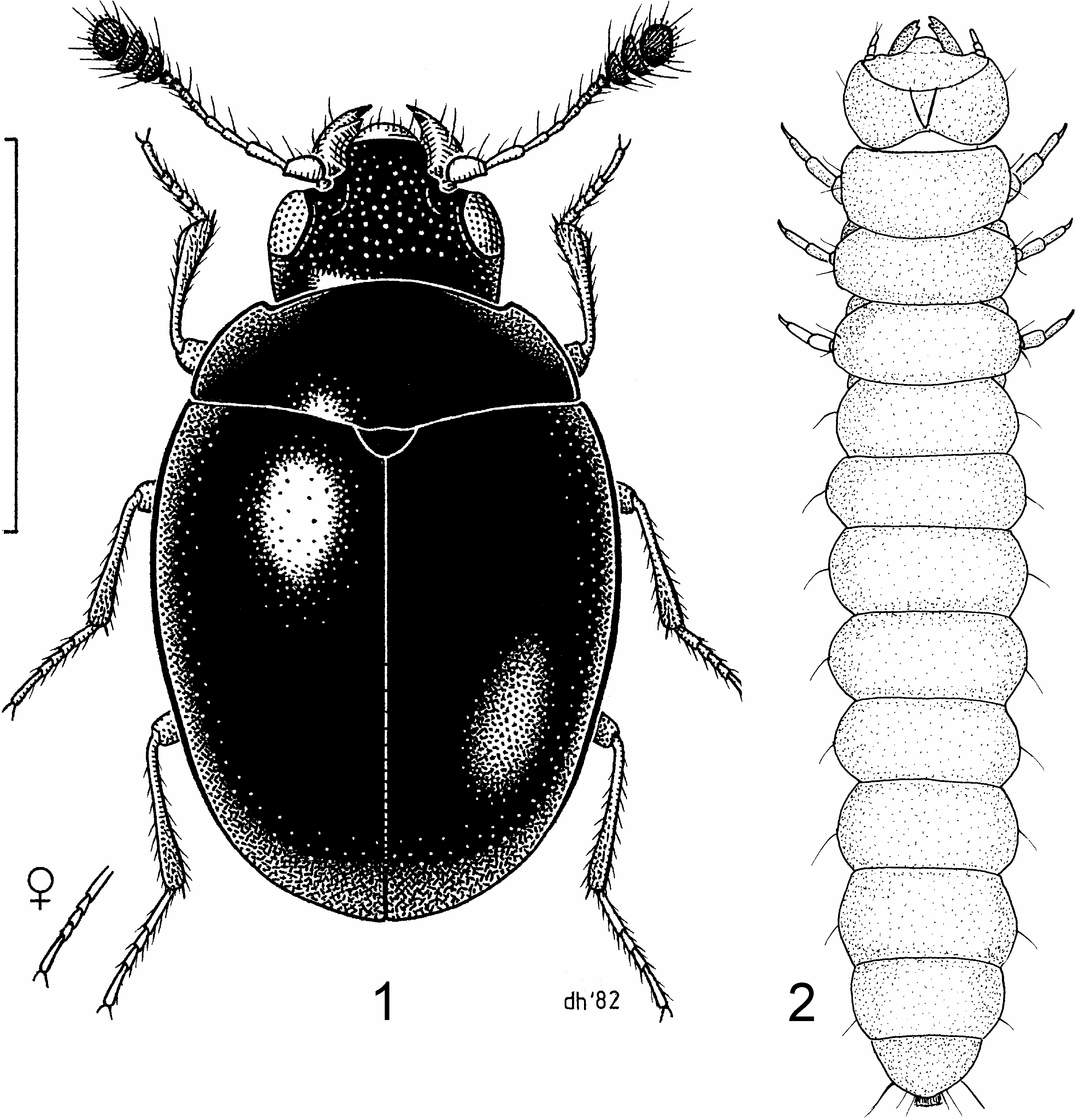
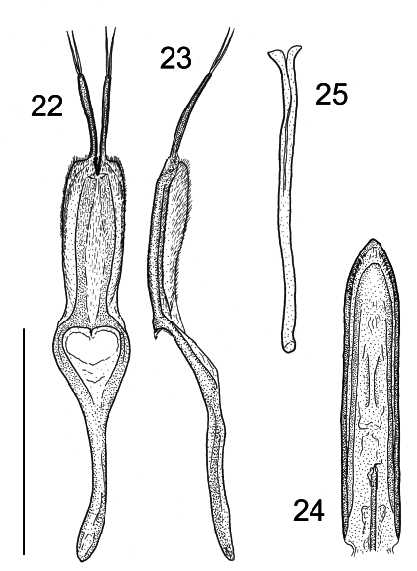
Adult description. Length 2.00– 2.76 mm. Body ( Fig. 1 View Figs ) 1.4–1.5 times as long as wide, nearly circular, very convex; dorsal surface smooth, shiny; vestiture of extremely sparse, inconspicuous, recumbent hairs. Head (Fig. 5) about 1.15 times wider than long, frons usually punctate, punctures coarse to fine ( Figs. 19–21 View Figs ); mouthparts anteriorly oriented; head not constricted behind eyes, transverse vertexal line absent; stridulatory file absent, temples about twice as long as eyes. Tentorial arms moderately separated at base, extending posteriorly from epistome to behind level of posterior margin of eye, slightly diverging, corpotentorial bridge narrow and median tendon absent. Frontoclypeal suture absent; clypeus extending well in front of antennal insertions, subrectangular at apex, sides arcuately emarginate above antennal insertions. Eyes moderately large, subcircular, prominent anteriorly, moderately coarsely facetted, without interfacetal setae. Antennae (Fig. 6) 11-segmented with a distinct 3-segmented club; antennal insertions exposed from above; genae with deep antennal grooves; genae not projecting. Gular sutures well separated. Cervical sclerites apparently absent. Labrum visible, broadly rounded. Mandible about 1.15 times as long as wide at base, bent abruptly mesally, with slight dorsal concavity, not setose; apex tridentate; mola well developed and transversely ridged; prostheca consisting of a wide membrane and brush of setae. Maxilla with galea 3–4 times as wide as lacinia; apex of lacinia with a pair of spines; terminal maxillary palpomere about 3 times as long as wide, widest just proximal to middle. Mentum transverse; apical labial palpomere about twice as long as wide, cylindrical. Pronotal length 0.40–0.56 mm, about 2.25 times wider than long, base as wide as elytral base; sides evenly arcuate, with distinct lateral bead; lateral carina sharp, complete, feebly explanate; anterior angles acute, protruding; anterior edge with margin, margin obsolete near midline; posterior edge strongly posteriorly arcuate, weakly margined; disc with sparse, irregularly spaced punctures. Prosternum (Fig. 7) short in front of coxae, about half as long as coxal cavity; male with medial rounded setiferous sex patch; prosternal process broad, not extended posteriorly beyond level of procoxae, sides expanded laterally at apex, apical edge straight; apicomedian process (not visible in ventral view) articulating with notch on mesoventrite. Notosternal suture complete. Procoxae not projecting. Procoxal cavities slightly transverse, with narrow lateral extension, externally broadly open behind, internally narrowly open behind; trochantins exposed. Mesoventrite short, convex, with vertical anterior procoxal rests. Mesocoxal cavities subcircular, separated by about their diameter, open laterally (partly closed by mesepimera), with exposed trochantins. Metaventrite about 1.5 times wider than long, convex, discrimen absent. Metepisternum long and narrow, about 6 times longer than wide. Metendosternite with broad, hyaline stalk, long arms, with long S-shaped ventrolateral projections arising from middle of arms. Metacoxae strongly transverse, subcontiguous, reaching metepisternum. Scutellum slightly wider than long, rounded posteriorly, almost semicircular, impunctate. Elytral length 1.22–1.70 mm, 1.72–1.74 times as long as wide and about 3 times as long as pronotum, humeri well developed, slightly obtuse; disc strongly and evenly convex, punctation very fine and sparse, punctures not in distinct rows. Epipleura (Fig. 8) complete, wide anteriorly, abruptly narrowed about midway to apex, with a deep longitudinal fovea in anterior half containing moderately dense, stout, erect setae. Hind wings reduced to short pads or fully developed; apical field more than 0.5 times wing length; radial cell present but reduced and somewhat oblique; linear sclerite present just beyond radial cell; r3 very short and r4 incomplete; basal portion of RP very short to moderately long, with vaguely indicated long apical extension; medial spur straight and not reaching wing margin; medial field with 4 free veins (sometimes with remnant of a fifth) and no medial fleck; wedge cell absent; anal notch deep. Legs moderately long, slender; trochanterofemoral joint oblique; femur inflated near middle; tibiae weakly compressed, slightly expanded at apex, not spined; tibial spurs weak, paired, equal in length. Tarsi 5-5- 5 in female and 5-5- 4 in male; penultimate tarsomere slightly lobed beneath; protarsomeres 1–4 subequal; mesotarsomere 1 about 1.5 times as long as 2, mesotarsomeres 2–4 subequal; metatarsomere 1 about twice as long as 2, penultimate and antepenultimate metatarsomeres subequal; pretarsal claws simple; empodium small, not projecting between claws. Abdomen with 5 free ventrites, ventrite I not much longer than II, without postcoxal lines; intercoxal process acute. Abdominal spiracles present on segments I–V. Sternite VIII in male without anterior strut, sternite IX in male apically membranous, basally with anterior strut (spiculum gastrale, Figs. 25 View Figs , 29 View Figs ).Aedeagus (Fig. 9) uninverted, with tegmen dorsal to penis; tegmen ( Figs. 22 View Figs , 26 View Figs ) with anterior strut; parameres free, contiguous at base. Penis ( Figs. 10, 11 View Figs ) with long basal strut, extending internally to prothorax when retracted; endophallus with long flagellum. Female sternite VIII with well-developed spiculum ventrale. Ovipositor ( Fig. 12 View Figs ) about 2 times as long as wide, with two pairs of elongate, slender baculi (dorsal and ventral); gonocoxite, valvifer, and paraproct subequal in length; paraproct with heavily sclerotised internal process on anterior edge; styli well developed, apically attached; spermatheca ( Fig. 13 View Figs ) sclerotised, C-shaped, duct with coils ( LAWRENCE et al. 1999b; LESCHEN et al., in press).
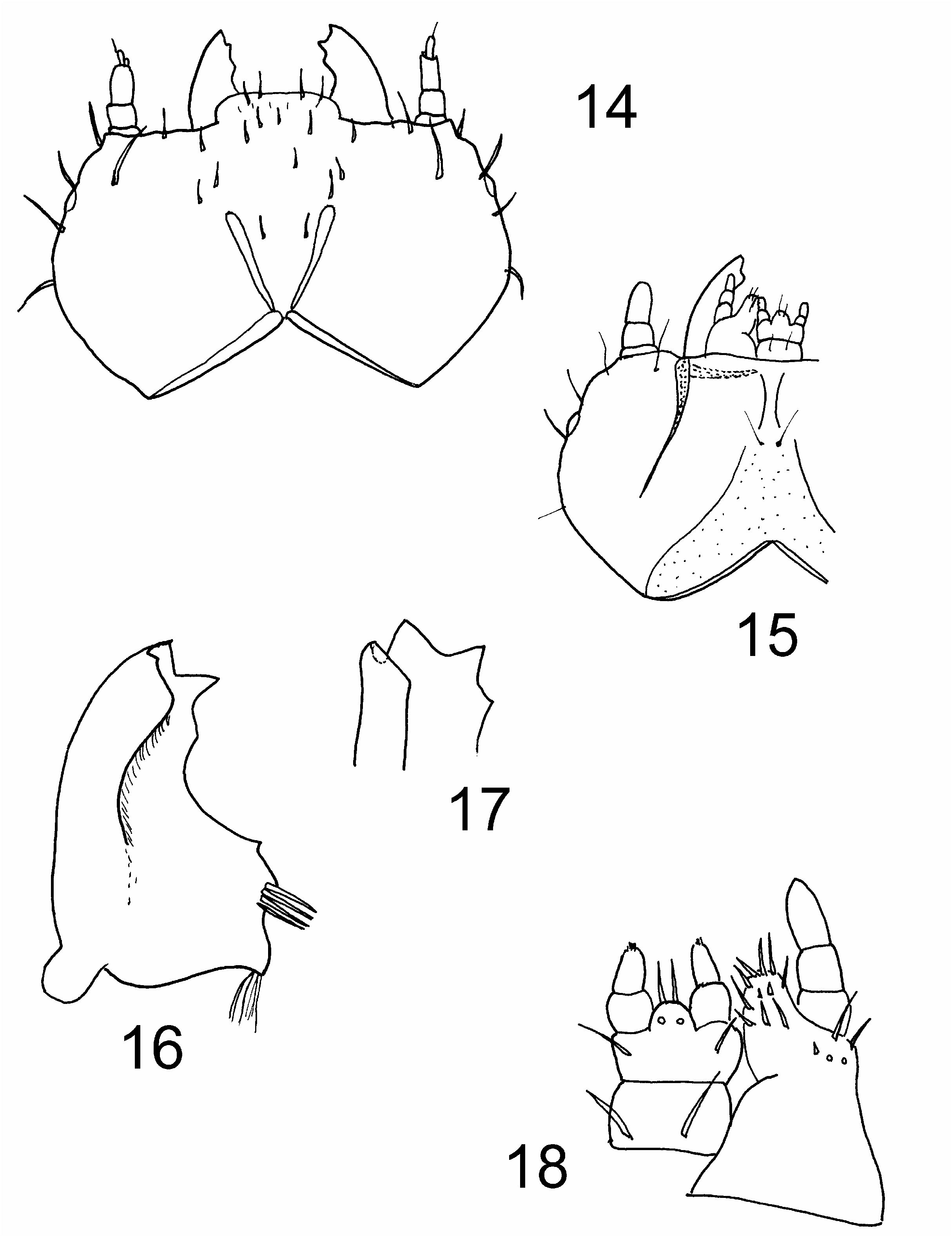
Larval description (based on C. politula ). Body ( Fig. 2 View Figs ) elongate, more or less parallel-sided and slightly flattened; very lightly pigmented, except for head capsule and tergum IX; dorsal surfaces smooth; vestiture consisting of fine hairs or setae. Head ( Fig. 14 View Figs ) prognathous, relatively transverse with strongly rounded sides; posterior edge of head capsule distinctly emarginate. Epicranial stem absent; frontal arms lyriform, contiguous at bases, initially Vshaped then suddenly angled and extending laterally to antennal sockets; frontal arms joined anteriorly by depigmented band; median endocarina absent; paired endocarinae present under V-shaped portion of frontal arms. Stemmata on each side 5. Antennae 3-segmented, less than 0.15 times as long as head width; first two antennomeres transverse, third elongate; sensorium shorter than apical antennomere, conical or palpiform. Frontoclypeal suture absent; labrum fused to clypeus, small, with long, widely separated tormae. Mandibles ( Figs. 16, 17 View Figs ) symmetrical, broad at base and narrow at apex, tridentate, without accessory ventral process; incisor edge with 1 or 2 subapical teeth; mesal surface of mandibular base with 2 to 5 hyaline processes, sometimes joined basally; mola absent. Ventral mouthparts ( Figs. 15, 18 View Figs ) protracted; cardo undivided, transverse; stipes wider than long; articulating area absent; mala simple, not cleft, rounded or truncate and setose or spinose; palps 3-segmented. Labium consisting of prementum and postmentum, which are basally connate with maxillae; ligula shorter than labial palp, simple; palps 2-segmented, separated by more than width of first palpomere. Hypopharyngeal sclerome absent. Hypostomal rods moderately long, subparallel and not extending to posterior of head; ventral epicranial ridges absent. Gular sutures very narrowly separate; gula longer than wide and not separated from labium by suture. Prothorax not longer than meso- and metathorax combined, terga with fine and sparse asperities, especially anteriorly; protergum without sclerotised plates. Prosternum with single, weakly defined sclerite. Legs well developed but relatively short, 5-segmented; pretarsus claw-like with 2 setae; mesocoxae separated by more than 2 basal coxal diameters. Abdomen more than twice as long as thorax; segments I–VII lacking ventrolateral processes, terga and sterna with lateral patches of fine asperities; paired abdominal glands absent. Abdominal segment IX shorter than VIII; tergum extending onto ventral surface but not forming articulated plate; urogomphi absent. Sternum IX simple, not enclosed by sternum VIII. Segment X without paired pygopods; anal region posteroventrally oriented. Spiracles annular-biforous, not placed at ends of spiracular tubes, those on segment VIII about the same size as others on abdomen, facing laterally; accessory chambers about as long as peritreme and facing posteriorly ( LAWRENCE et al. 1999a; LESCHEN et al., in press).
Differential diagnosis. Adults of this family may be distinguished from all other Coleoptera by the highly convex body form, eighth antennomere not significantly reduced in size, ventral antennal grooves present on the head, externally broadly open procoxal cavities, deep elytral epipleural foveae lined with stiff setae, and 5-5-4 tarsal formula in the male. Adults can be further separated from similar-looking Phalacridae by the antennal insertion exposed, lacinial uncus absent, protrochantin exposed, elytral punctation confused and not seriate, mesocoxal cavity open laterally, and functional spiracles absent on abdominal segment VII. Among Cucujoidea the larvae of Cyclaxyridae are distinguished by their parallel body without pigmented terga, head capsule posteriorly emarginated, paired endocarinae contiguous at base and anteriorly divergent, lack of urogomphi, protracted ventral mouthparts, mesal surface of mandibular base with hyaline process, inner apical angle of mala rounded or truncate, and labial palpi 2-segmented, pretarsus bisetose, segment X and anal opening posteroventral, and spiracles annular-biforous.
Composition and distribution. The family contains only the genus Cyclaxyra , with two species (one newly described below) occurring on the North, South, and Stewart Islands in New Zealand. The two species have not been taken at the same site, and C. politula appears to be mostly absent from northeastern South Island. A Quaternary fossil identified as C. impressa Broun, 1915 has been reported by MARRA et al. (2008) from Taranaki.
Phylogenetic relationships. In a phylogenetic analysis of 99 larval and adult characters, Cyclaxyridae was placed as sister taxon to the Australian family Tasmosalpingidae by LESCHEN et al. (2005), a placement that was supported mainly by adult characters, as this sister-relationship is shown by adult-only trees. Possibly, Cyclaxyra is more closely related to Lamingtoniidae ( LAWRENCE & LESCHEN 2003) , as suggested in larval-only trees.
Biology. Both larvae and adults are inhabitants of sooty-mould fungi (Ascomycota: Dothideomycetes: Capnodiales), which in south temperate regions form thick black “wefts” of mycelia of up to six different species ( HUGHES 1972: 226), generally on the surfaces of foliage, branches, tree trunks, and often surrounding rocks and soil. These growths are associated with sugary exudates of leaves or with the “honeydew” produced by sternorrhynchous Hemiptera, primarily in association with Nothofagus , but other plants as well ( KLIMASZEWSKI & WATT 1997, CARLTON & LESCHEN 2007). Based on examination of gut contents the beetles feed on spores, conidia, and hyphae of the fungus. This habit has evolved in multiple lineages of Coleoptera , and in New Zealand and elsewhere beetles in several genera and families are also associated with sooty moulds (e.g., Nothoderodontus Crowson, 1959 ( Derodontidae ; found in temperate South America, New Zealand and Australia), Metaxina Broun, 1909 ( Metaxinidae ; family endemic to New Zealand), Agapytho Broun, 1921 ( Agapythidae ; family endemic to New Zealand), Hisparonia Kirejtshuk, 2004 ( Nitidulidae ; genus endemic to New Zealand), Triphyllus Dejean, 1821 ( Mycetophagidae ; one species associated mainly with sooty moulds), and Doxozilora Broun, 1909 ( Melandryidae ; genus is endemic to New Zealand); see JOHNSON et al. 2008).
Cyclaxyra species are active at night and may be collected in large numbers on trunks of trees and larger shrubs where sooty moulds are growing. Mating has also been observed at this time where males mount females on top (not back to back) (LESCHEN, pers. observ.).
The function of the epipleural foveae remains unknown. We have not observed spores within these, and the structure is not sexually dimorphic.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Cyclaxyridae
| Gimmel, Matthew L., Leschen, Richard A. B., Ślipiński, S. Adam & Csiro 2009 |
Cyclaxyrinae
| LESCHEN R. A. B. & LAWRENCE J. F. & SLIPINSKI S. A. 2005: 63 |
| LAWRENCE J. F. & LESCHEN R. A. B. 2003: 910 |
| KLIMASZEWSKI J. & WATT J. C. 1997: 49 |