
Trichomycterus rubbioli, Bichuette, Maria Elina & Rizzato, Pedro Pereira, 2012
|
publication ID |
https://doi.org/ 10.5281/zenodo.282362 |
|
DOI |
https://doi.org/10.5281/zenodo.6175196 |
|
persistent identifier |
https://treatment.plazi.org/id/03D4C111-6F08-FFA7-FF0E-FF0E7B87F3C3 |
|
treatment provided by |
Plazi |
|
scientific name |
Trichomycterus rubbioli |
| status |
sp. nov. |
Trichomycterus rubbioli View in CoL sp.n.
( Fig. 2 View FIGURE 2 )
Trichomycterus sp. 3— Trajano (2003).
Trichomycterus undescribed species—Trajano et al. (2009).
Trichomycterus undescribed species 2— Proudlove (2010).
Trichomycterus View in CoL sp. 2— Trajano & Bichuette (2004); Mattox et al. (2008); Trajano & Bichuette, (2010); Trajano et al. (2011).
Holotype. MZUSP (110977), 60.3 mm SL, Brazil, south-western State of Bahia, Carinhanha County, Lapa dos Peixes cave (13° 49’ 21,78”S 43° 57’ 24,39”W), Middle São Francisco River basin, Serra do Ramalho karstic area, 23 April 2000, M. E. Bichuette, E. Trajano & D. Möeller.
Paratypes. All from Brazil, southwestern State of Bahia, Carinhanha County, Middle São Francisco River basin, Serra do Ramalho karstic area. MZUSP (110978), 1, 56.8 mm SL, Lapa dos Peixes cave, 23 April 2000, M. E. Bichuette, E. Trajano e D. Möeller; MZUSP (110979), 1, 64.1 mm SL, Lapa dos Peixes cave, 23 April 2000, M. E. Bichuette, E. Trajano & D. Möeller; MZUSP (110980), 1, 32.9 mm SL (C&S), Lapa dos Peixes cave, 23 April 2000, M. E. Bichuette, E. Trajano & D. Möeller; MZUSP (110981), 1, 29.7 mm SL, Gruna da Água Clara cave (13° 48’ 2.83” S 43° 57’ 4.76” W), 11 July 1999, L. Senna-Horta; MZUSP (110982), 1, 29.9 mm SL, Gruna da Água Clara cave, 11 July 1999, L. Senna-Horta; MZUSP (110983), 1, 58.6 mm SL, Gruna da Água Clara cave, 11 July 1999, L. Senna-Horta; MZUSP (110984), 1, 31.3 mm SL, Gruna da Água Clara cave, 22 April 2000, M. E. Bichuette & E. Trajano; MZUSP (110985), 1, 21.2 mm SL, Gruna da Água Clara cave, 22 April 2000, M. E. Bichuette & E. Trajano; MZUSP (110986), 1, 16.9 mm SL, Gruna da Água Clara cave, 22 April 2000, M. E. Bichuette & E. Trajano.
Diagnosis. Trichomycterus rubbioli is readily distinguished from epigean and most hypogean congeners by three distinctive, non-exclusive characters: highly reduced skin pigmentation (except for T. gorgona and caverestricted congeners, T. chaberti, T. itacarambiensis , T. spelaeus, T. sandovali, T. santanderensis, T. uisae and T. dali ) ( Fig. 2 View FIGURE 2 ); reduced eyes, visible externally as small black or reddish-brown spots (except for T. gorgona and the hypogean T. chaberti, T. itacarambiensis , T. santanderensis, T. uisae and T. sketi) ( Figs. 2 View FIGURE 2 and 3 View FIGURE 3 ); and barbels long, especially nasal (86.9–127.1% of HL) and maxillary (98.1–129.3% of HL) (except for T. longibarbatus and the hypogean T. spelaeus, T. santanderensis, T. uisae, T. sketi and T. dali ) ( Figs. 2 View FIGURE 2 and 3 View FIGURE 3 ). The characters above, in combination, differentiate new species from all hypogean congeners except for T. santanderensis and T. uisae. Trichomycterus rubbioli can be further diagnosed from almost all congeners, including T. santanderensis and T. uisae (except for T. hualco, T. sketi and T. dali ) by pectoral-fin ray count reaching I, 9.
Description. Morphometric data of holotype and paratypes are given in Table 1.
Holotype Range (n=8) Mean SD Standard length 60.3 14.4–64.1 40.4 19.4 Percents of standard length
Total length 119.3 112.3–121.0 117.0 2.7 Predorsal length 60.9 57.3–60.9 59.3 1.2 Preanal length 76.7 70.5–78.2 74.3 2.6 Prepelvic length 65.1 59.3–65.3 62.2 2.3 Caudal peduncle length 26.2 14.3–26.3 21.7 4.1 Caudal peduncle depth 16.3 8.1–16.3 13.9 2.8 Dorsal-fin base length 10.8 8.8–15.0 11.3 1.7 Anal-fin base length 10.7 7.8–11.4 9.3 1.1 Pectoral-fin base length 5.1 4.9–6.4 5.5 0.4 Pelvic-fin base length 3.2 2.5–3.2 2.7 0.2 Dorsal-fin length 17.4 15.1–22.5 18.3 2.4 Anal-fin length 14.6 14.2–19.6 16.0 2.1 Pectoral-fin length 23.3 19.1–23.6 22.4 1.4 Pelvic-fin length 10.2 8.7–12.4 10.7 1.2 Head length 22.2 19.8–23.2 21.8 1.3 Body depth 12.7 11.4–13.3 12.4 0.6 Body width 19.4 18.1–22.4 19.9 1.4 Percents of Head length
Head depth 87.2 83.2–98.3 91.8 5.0 Head width 57.5 50.4–63.8 57.2 4.5 Interorbital width 34.1 27.7–34.9 32.2 2.3 Eye diameter 7.3 4.0–7.3 6.1 1.1 Snout length 15.4 13.9–24.0 19.1 4.2 Maxillary barbel length 129.3 98.1–129.3 109.9 9.7 Rictal barbel length 77.1 50.5–77.1 60.1 9.5 Nasal barbell length 86.9 86.9–127.1 103.3 15.4 Mouth width 70.7 28.5–70.7 49.1 16.3 Body elongate semi-cylindrical, becoming compressed towards the caudal fin ( Fig. 2 View FIGURE 2 ). Dorsal profile of the body straight, very slim, with a straight slope from the tip of the snout to the anterior portion of trunk; ventral profile of body straight in lateral view. Dorsal profile of caudal peduncle slightly convex to beginning of caudal fin; ventral portion of caudal peduncle straight. Caudal fin spatulate, with distal margin straight. Dorsal-fin origin on posterior half of body, slightly anterior to origin of pelvic fin. Anal-fin origin posterior to distal margin of dorsal fin. Adipose cutaneous folds present in young and adult specimens along the dorsal profile of the body: anterior to dorsal fin (pre-dorsal) well developed in young, poorly developed to absent in adult specimens, not supported by procurrent rays; posterior to dorsal fin (post-dorsal) well developed in young and relatively reduced in adult specimens, slightly convex in lateral view, supported by procurrent rays only in the posterior end. Urogenital and anal openings on vertical through posterior end of dorsal-fin base, between distal margins of pelvic fins.
Head relatively wide and depressed, semicircular in adult and trapezoidal in young specimens in dorsal view ( Fig. 3 View FIGURE 3 ). Adductor mandibulae muscles well developed in adults, easily noticeable externally, dorsally delimited by a longitudinal facial groove. Very reduced eyes, visible externally as small black or reddish-brown dots. Eyes more superficial especially in some smaller individuals, covered with a thin layer of skin and with very regular forms (rounded); others, especially larger individuals, with deeper eyes (covered by a thick layer of skin), with irregular form. On at least one individual ( MZUSP, 110983), right eye not visible externally; holotype with pale red eyes. Anterior nostril transversally ovoid and slightly smaller than posterior, surrounded laterally by nasal barbels. Posterior nostril rounded, surrounded anteriorly by large flap of integument. Mouth slightly subterminal, convex in dorsal view, rictus laterally directed. Barbels long, especially nasal and maxillary. Maxillary barbel, extending to middle of pectoral fin; nasal barbel extending to origin of pectoral fin, and submaxilary barbel extending to opercle. Opercular patch of odontodes very small and circular. Interopercular odontodes forming a slightly convex patch throughout ventral margin of interopercle. Opercle with 5–13 odontodes, interopercle with 35–38 odontodes.
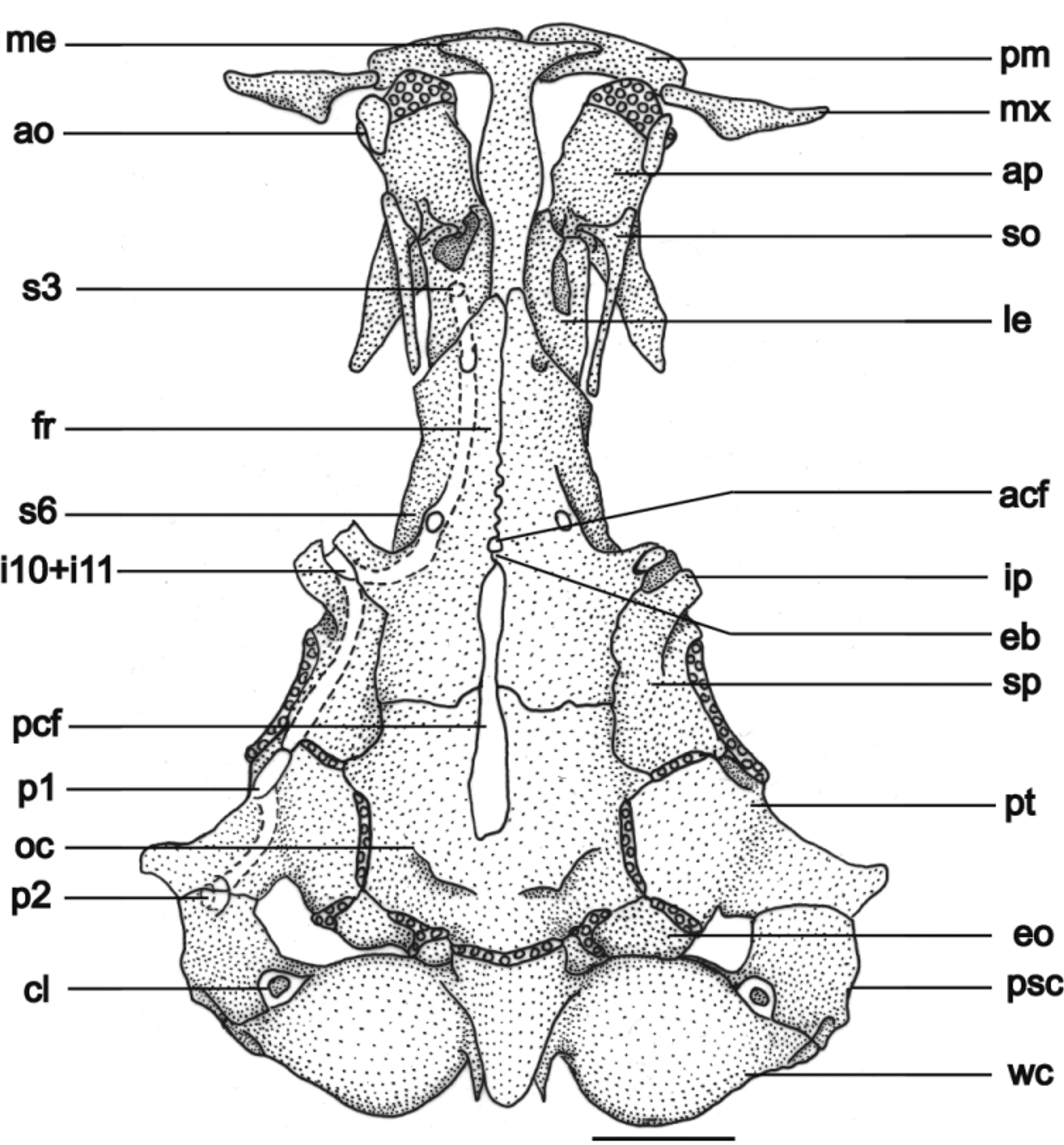
General morphology of cranium. Cranial fontanels divided by an epiphyseal bar. Anterior fontanel small and rounded, almost at middle length of the frontal bones; posterior fontanel extending from the middle of parietosupraoccipital to the posterior region of the frontal bones, with a conspicuous constriction at intersection of parietosupraoccipital and frontal bones ( Fig. 4 View FIGURE 4 ). Parieto-supraoccipital with two posterodorsal processes as insertions to the epiaxialis and supracranialis anterior muscles. Lateral process of pterotic pointing diagonally outwards, in direction to opercular patch of odontodes. Anterior process of sphenotic and posterolateral process of frontal conspicuous, hollow horn-like structure pointing diagonally forward, inside of which emerge infraorbital sensory branches.
Sesamoid-supraorbital long and cylindrical, with internal, anterolateral projection, pointing to mesethmoid. This projection may be long and sharp or small and slightly convex. Autopalatine with straight anterior margin covered with cartilage, almost rectangular medial region and long posterior process that becomes sharp on distal region and totally covers the internal half of metapterygoid. Distal profile of mesethmoid straight in dorsal view, main body axis with enlarged medial region, cornua reaching half of premaxillary length. Vomer arrow-shaped with long posterior process wider at middle length. Parasphenoid almost circular at medial region, with long and narrow posterior process reaching Weberian complex and two anterior processes surrounding laterally the posterior process of vomer. Three to five irregular premaxillary rows of conic teeth curved backwards. Maxilla boomerang shaped, rounded proximal margin of maxilla joining premaxilla, anterior to antorbital. Lower jaw with conical teeth, curved backwards; three to four rows with larger teeth at middle and one row externally, close to upper process of dentary, with teeth gradually becoming smaller. Hyomandibular with a conspicuous semicircular depression, joined tightly to metapterygoid and quadrate ( Fig. 5 View FIGURE 5 ). Metapterygoid and quadrate united together by anterior block of cartilage. Opercle with dorsal process on which inserts levator operculi and dilatator operculi muscles, with reduced patch of odontodes. Preopercle with sharp posterior region, articulating to dorsal process of interopercle by straight ventral margin, not covered by cartilage. Interopercle long and wide, with dorsal, T-shaped process articulating to preopercle; anterior process without odontodes, bordered ventrally and posteriorly by long, sharp odontodes.
Branchial skeleton and associated structures. Branchiostegal-rays seven. Rays 5, 6 and 7 with enlarged distal extremity. Ray 7 covered by interopercle, reaching ventral margin of opercular patch of odontodes. Urohyal with long, very narrow dorsal process almost reaching cleithrum, broad convex posterior margin, urohyal-foramen slightly ovoid, anterior process T-shaped. Ventral hypohyal with depression on which articulates anterior process of urohyal ( Fig. 6 View FIGURE 6 ). Basibranchials 3, hypobranchials 3, ceratobranchials 5, epibranchials 5, pharyngobranchials 2 ( Fig. 7 View FIGURE 7 ). Basibranchial 1 absent. Basibranchial 2 and 3 connected to each other by their cartilaginous tips, forming long rod. Basibranchial 2 relatively wide with convex anterior margin. Basibranchial 3 with rectangular, rod-like posterior half, divergently triangular on anterior half, with convex anterior margin. Posterior tip of basibranchial 3 covered by cartilaginous posterior portion of hypobranchials 3. Basibranchial 4 cartilaginous, approximately hexagonally shaped. Anterior margins of basibranchial 4 bordered by cartilaginous posterior portion of hypobranchials 3, lateral and posterior margins bordered by cartilaginous anterior tips of ceratobranchial 4 and ceratobranchial 5, respectively. Hypobranchial 1 with external portion wider than internal, with cartilaginous tips. Hypobranchial 2 boomerang shaped, posterior half cartilaginous, anterior half ossified, forming anterior process that almost reaches external posterior margin of hipobranchial 1. Hypobranchial 3 almost completely cartilaginous, only anterior tip ossified and triangular, closely 88joined to anterior cartilaginous tip of ceratobranchial 3. Hypobranchial 4 absent. Ceratobranchials slightly curved, with cartilaginous extremities. Ceratobranchial 1 with medial tip wider than distal. Ceratobranchial 2 with shallow concavity on its posterior margin, without defined posterior process. Ceratobranchial 3 with pronounced concavity on its posterior margin, limited posteriorly by very small process. Ceratobranchial 5 slightly enlarged, bearing patch of small, narrow conical teeth pointed dorsally on its anterior half, connected to epibranchial 5 only by upper half of posterior margin. Posterior margin of ceratobranchials 3, both margins of ceratobranchials 4 and anterior margin of ceratobranchials 5 bearing one row of conical, very sharp teeth (not shown in fig. 7). Epibranchials 1, 2 and 3 narrow, rod-like, with cartilaginous tips. Epibranchial 1 with long, narrow and sharp anterior process, pointed outwards in acute angle. Epibranchial 2 with two small, acute processes, not uncinate, at the middle of anterior margin and at the external portion of posterior margin. Epibranchial 3 with conspicuous, posteriorly directed, large uncinate process at external portion of its posterior margin. Epibranchial 4 large, curved, with a wide, straight dorsal margin joined to posterior half of tooth plate and covered with cartilage, and with ventral margin very narrow, ovoid and cartilaginous. Epibranchial 5 very small, curved and completely cartilaginous, joining the posterior cartilaginous tip of ceratobranchial 4 with the ventral margin of epibranchial 4. Pharyngobranchials 1 and 2 absent. Pharyngobranchial 3 elongate, rod-like, slightly depressed, with cartilaginous tips. Pharyngobranchial 4 cartilaginous, curved, joined tightly to dorsanterior half of tooth plate. Tooth plate well developed, curved, with two irregular rows of long, conic, internally curved teeth; internal row with thicker, robust teeth.
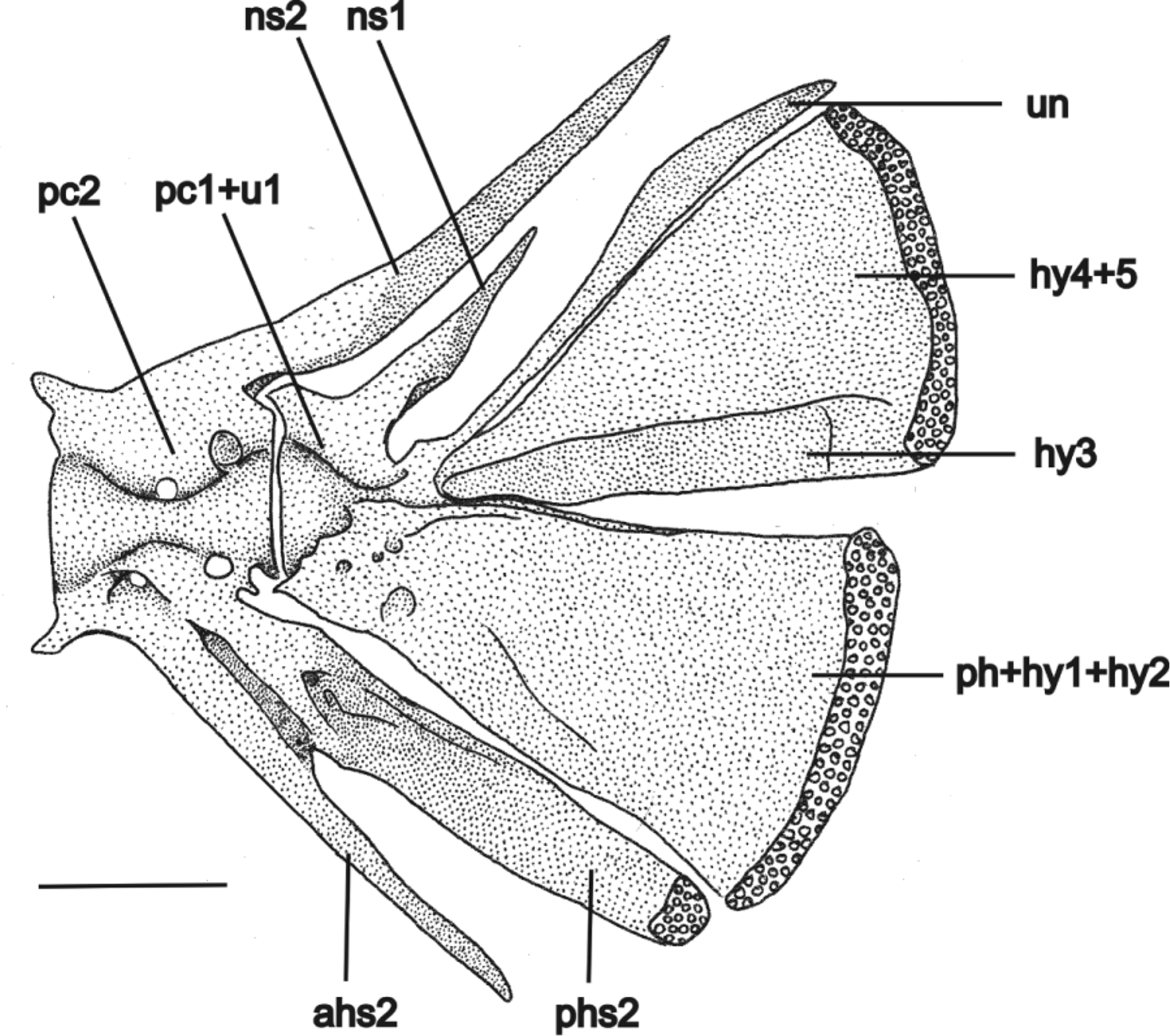
Postcranial skeleton. Total vertebrae 31–32, 13–15 pairs of ribs. First rib thicker, posteriormost gradually thinner. Pectoral girdle with wide cleithrum, becoming narrower at anterior half, joined together by very narrow anterolateral margin. Posterior process of pectoral girdle reaching ventral surface of posttemporo-supracleithrum. Scapulocoracoid with conspicuous anterior process projected forward, with narrow base and wide apex reaching external margin of cleithrum. Proximal radial 1 cartilaginous, joined to first and second pectoral-fin rays. Proximal radial 2 cartilaginous, small. Proximal radial 3 large, well ossified, with semicircular cartilaginous posterior margin joined to third to tenth pectoral-fin rays, and small, ovoid anterior cartilaginous margin, joined to scapulocoracoid ( Fig. 8 View FIGURE 8 ). Pelvic girdle with external anterior process narrower than internal anterior process. Short medial process sometimes present, pointing forward ( Fig. 9 View FIGURE 9 ). Epural absent. Neural spine of compound caudal centrum reduced to half or one-fourth of uroneural length, sharp tip projected upward. Hemal spine (n = 2) of penultimate vertebrae divided in a narrow anterior hemal spine and a wide posterior hemal spine with cartilaginous tip bearing one procurrent ray. Parhypural fused to hypural 1+2 into trapezoidal lower hypural plate. Hypural 3 partially fused to hypural 4+5, into triangular upper hypural plate. Uroneural not fused to hypural 4+5, long, reaching distal margin of upper hypural plate, with characteristic shape: proximal half narrow and distal half expanding and sharpening again distally, similar to a scimitar blade ( Fig. 10 View FIGURE 10 ).
Pectoral fin triangular in dorsal view, with wide base and first ray long, thick, unbranched and filamentous. Pectoral-fin ray count I, 8 or I, 9, even at same individual. Dorsal fin approximately semicircular in lateral view, with nine rays, two first unbranched (II, 7); in three paratyes, including one C&S, I, 7; one or two micro rays anterior to first ray. Pterygiophores with seven or eight narrow basal radials in dorsal fin, slightly curved on basal portion, with spatulate distal region curved backwards and cartilaginous distal tip. First basal radial with narrow cartilaginous distal margin pointing backwards. Last basal radial with laminar posterior expansion on distal fold, joined to two last dorsal fin rays. Each basal radial, except for first and two last, joined to small distal radial, between two basal branches of each ray. First distal radial larger than posterior radials. Anal fin approximately rectangular in lateral view, distal margin rounded, seven rays (II, 5), two micro rays anterior to first ray. On two specimens, I, 6. Pterygiophores with six narrow basal radials on anal fin, morphologically similar to dorsal pterygiophores: spatulate, curved distal region, with cartilaginous tips, last with laminar posterior expansion articulating with the two last rays, each basal radial except first and two last joined to small distal radial ossified between two basal branches of each ray. Pelvic fin rectangular in ventral view, separated from each other, I, 4, one pelvic splint. Caudal fin with 14 principal rays (six in upper lobe, first unbranched; seven in lower lobe, first unbranched), 8–14 dorsal procurrent rays; 5–11 ventral procurrent rays.
Color in alcohol. Body generally pale yellow, resembling faded irregular spots. Dorsal fold (anterior and posterior to dorsal fin) lighter than body. Mouth, dorsal region of the head between eyes, and barbels light yellow to white. Eyes, when visible, black or dark brown, with one individual with red eyes. Pectoral, dorsal, pelvic, anal and caudal fins translucent ( Fig. 2 View FIGURE 2 ).
Live color. Body unpigmented. Internal organs can be seen by transparence. When subject to illumination for several hours, as during chronobiological experiments (12:12 light (1000 lux): dark cycles, during seven days; Trajano et al., 2011), the fish became very dark, due to light-induced synthesis of melanin. After several weeks, specimens of T. rubbioli became pale again, and the progressive loss of melanin followed a pattern of large, irregular spots, different from the natural color pattern.
Etymology. The specific name (noun in apposition) is in honor of Ezio Rubbioli , speleologist, the first explorer of Serra do Ramalho caves, who brought this new species to our attention.
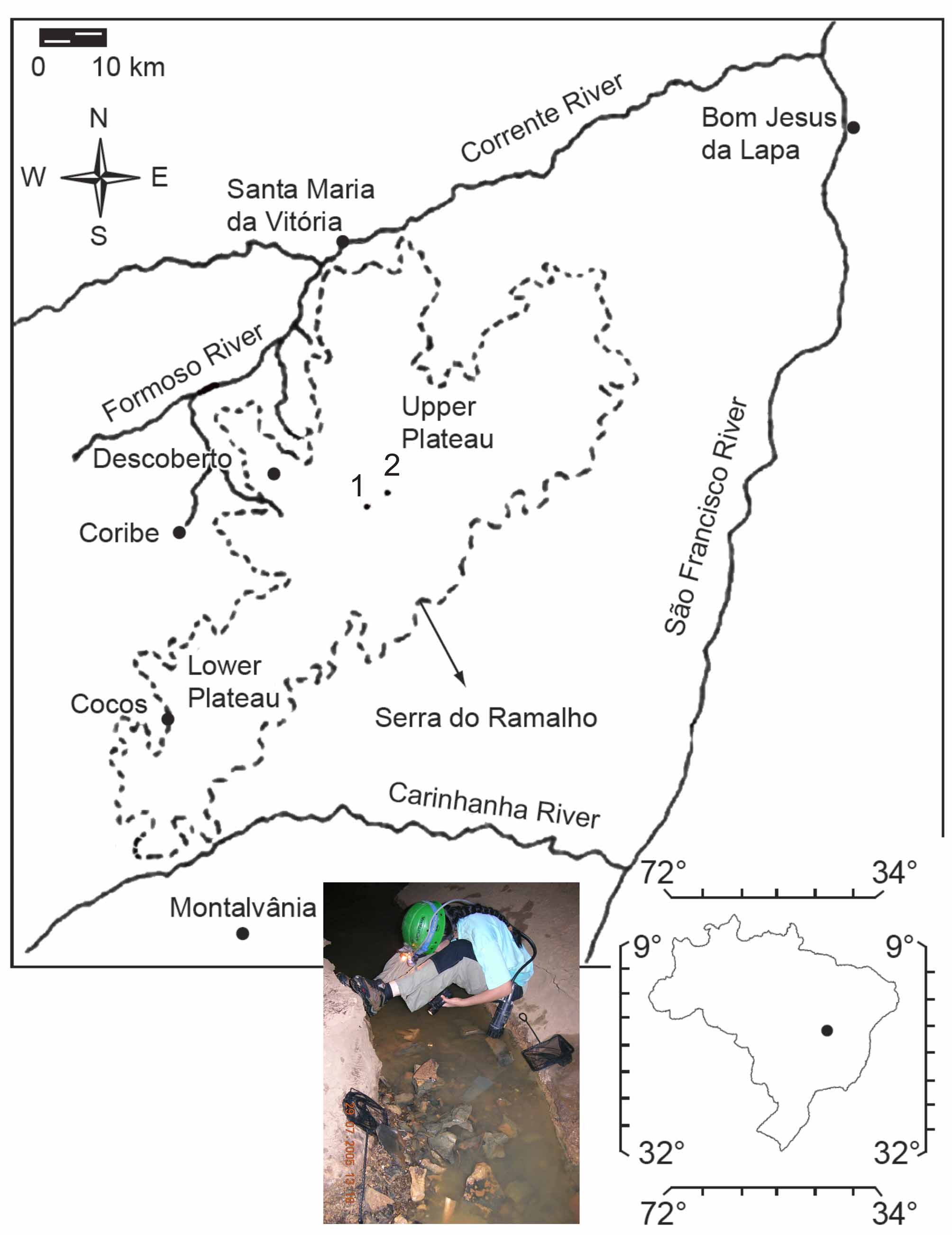
Distribution. Trichomycterus rubbioli is known exclusively from subterranean waters in at least two caves of the same cave system in the Serra do Ramalho karst area, middle São Francisco River basin: Lapa dos Peixes and Gruna da Água Clara ( Fig. 1 View FIGURE 1 ).
Notes on habitat and ecology. The small vadose stream inside Lapa dos Peixes where T. rubbioli catfishes were found is about 180 m long (accessible part), with an average width of 0.3 m and maximum depths of 0.5 m. The fishes were concentrated in places with the bottom formed by clay and small blocks of limestone, usually hiding under these blocks and inside crevices in the walls, thus showing a strong cryptobiotic habit. The level of water in this stream is lower in the dry season, and on two occasions the locality was mostly dry. Non-troglobitic fish, such as tetra characins, Astyanax sp., and heptapterids, Rhamdia sp. and Pimelodella sp., were observed in the locality ( Trajano et al., 2009). In April 2000 and September 2003, respectively, 15 and 17 specimens of T. rubbioli were counted along the 180 m long stream, but afterwards this number sharply decreased (see Trajano et al., 2009: Table 1), which suggests that the population in the vadose stream inside Lapa dos Peixes is a sink population, i.e., if the source of migrants is cut-off it can become extinct ( Fong, 2004).
When not hidden, T. rubbioli catfishes are fast swimmers, showing preference for staying on the bottom and on walls (rocky and silt substrate), with occasional incursions into the midwater and surface. The light from spotlights, 1.000 lux (light intensity of a cloudy day) triggers a quick avoidance reaction. In some occasions, the fishes were observed digging the soft bottom, apparently in a foraging behavior. Chronobiological studies indicated a partial regression, not as advanced as in highly troglomorphic species, of the circadian mechanisms of time control ( Trajano et al., 2009).
Some adult specimens collected in 2000 were maintained alive until 2008 in captivity, indicating a long lifespan.







| MZUSP |
Museu de Zoologia da Universidade de Sao Paulo |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.