
Paragrillotia spratti ( Campbell & Beveridge, 1993 ) Beveridge & Justine, 2007
|
publication ID |
https://doi.org/ 10.5281/zenodo.5390741 |
|
publication LSID |
lsid:zoobank.org:pub:2EBB7E08-C2D5-4190-91D3-C9F243453E26 |
|
persistent identifier |
https://treatment.plazi.org/id/03D65D17-FF81-FF99-113F-6F74FDB5FBE6 |
|
treatment provided by |
Marcus |
|
scientific name |
Paragrillotia spratti ( Campbell & Beveridge, 1993 ) |
| status |
comb. nov. |
Paragrillotia spratti ( Campbell & Beveridge, 1993) View in CoL n. comb.
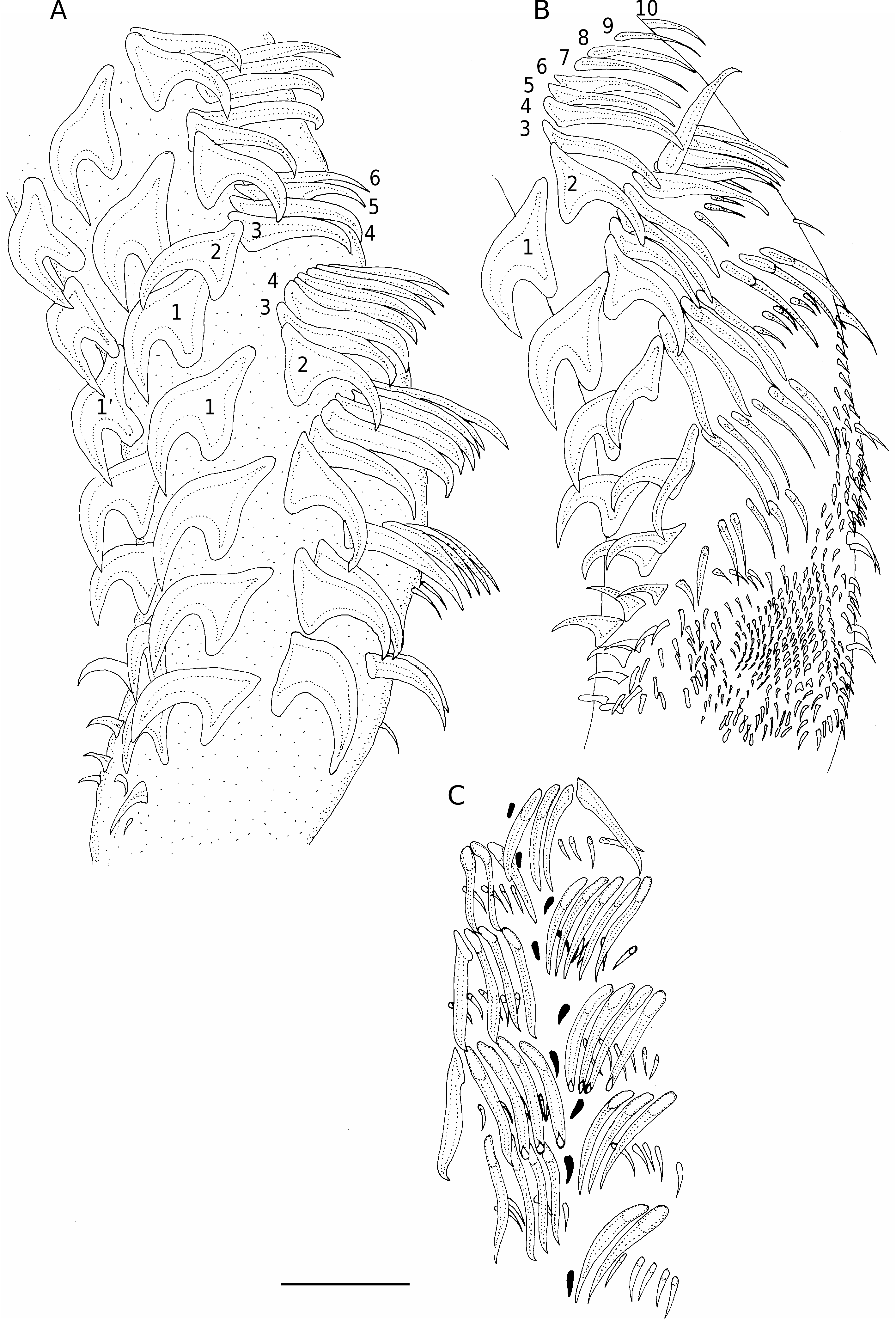
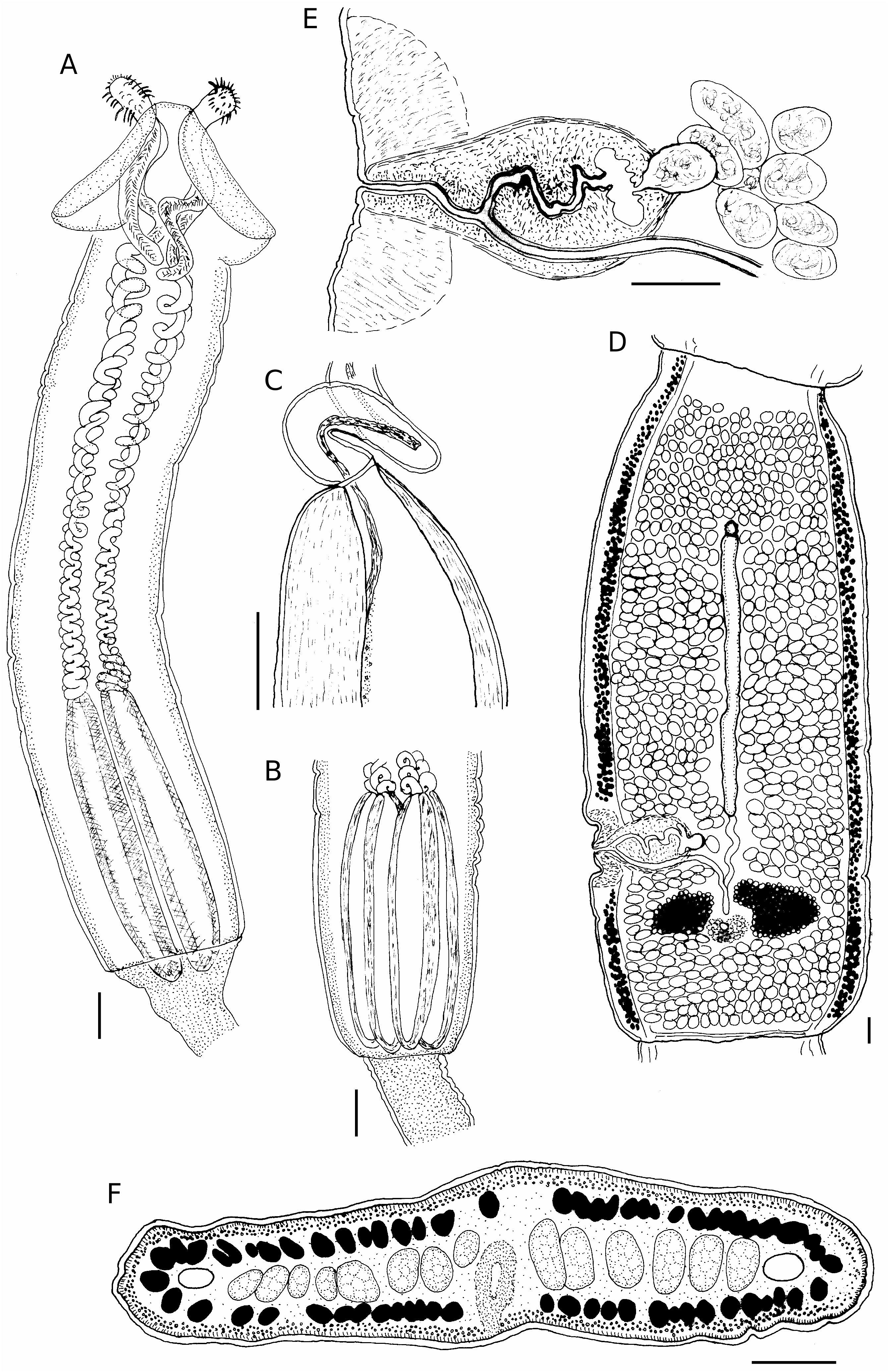
( Figs 3 View FIG ; 4 View FIG )
Pseudogrillotia spratti Campbell & Beveridge, 1993: 41-43 View in CoL , figs 10-16.
MATERIAL EXAMINED. — Paratypes from spiral valve of Negaprion acutidens , off Nouméa , New Caledonia, 15.V.2003, coll. J.-L. Justine, 18 specimens ( SAM 23331) (balsam mounts on 17 slides, MNHN JNC 441 E1 View Materials - JNC 441 E17; tentacles mounted in glycerine jelly on 5 slides
MNHN JNC 441E18- JNC 441E22; serial sections on 19 slides JNC 441E23- JNC 441E41).
DESCRIPTION
Longest specimen 93 mm long, maximum width 1300, with 110 segments, terminal segments mature. Scolex either slightly craspedote or acraspedote, 4.45-6.00 (4.96, n = 10) mm long, maximum width in region of pars bulbosa 850-1100 (970, n = 10); 2 oval bothria with median indentation in posterior margin, pars bothrialis 800-950 (890, n = 10); pars vaginalis 2700-4270 (3240, n = 10), sheaths spiral; bulbs 1500-1820 (1680, n = 10) long, 250-320 (290, n = 10) wide; bulb length:width ratio 1:5.2-7.2 (1:5.9, n = 10); prebulbar organ absent; retractor muscle originates in anterior region of bulb, insertion continued posteriorly by band of tissue on internal surface of bulb; pars post bulbosa absent, bulbs frequently project into pars proliferans scolecis. Mean scolex ratio 1:3.60:1.89.
Tentacles incompletely everted, maximum length 700; tentacle diameter 130-180 (150, n = 10) at base; no basal swelling.Tentacular armature heteroacanthous, heteromorphous; hooks hollow; principal rows of 9 or 10 hooks beginning on internal surface of tentacle, terminating on external surface; space present between hook files 1(1’) on internal surface. Hooks 1(1’) large, uncinate, with broad base, 113-135 (128, n = 5) long, base 83-98 (93, n = 5) long; hooks 2(2’) uncinate, smaller, more erect with shorter base, 100-113 (108, n = 5) long, base 65-70 (68, n = 5) long; hooks 3(3’) erect, falcate, 98-110 (105, n = 5) long, base 30-40 (35, n = 5) long; hooks 4(4’) to 10(10’) slender, erect with recurved tip, short base, gradually diminishing in size along row; hooks 4(4’) 108-118 (113, n = 5) long, base 23-35 (30, n = 5) long, hooks 10(10’) 80-98 (89, n = 5) long, base 20-43 (30, n = 5) long. Single row of slender 4- 7 intercalary hooks, 20-35 (26, n = 5) long, base 5-8 (6, n = 5) long; intercalary hook row begins posterior to hooks 6(6’), terminates between rows 9(9’) and 10(10’). External surface of tentacle with chainette of tiny hooks, one chainette element per principal row; chainette hooks at end of principal rows; chainette hooks uncinate, 13-18 (14, n = 5) long. No distinctive basal armature on internal surface; disparity between sizes of hooks 1 and 1’; sizes of hooks in ascending series of file 1 from base 70, 73, 100, 108, 115; comparable hooks in file 1’ 110, 120, 138, 138, 133 long. On external surface of tentacle, initial rows of small hooks blunt-tipped, 8-18 (14, n = 5) long; anterior to blunt hooks, compact triangular array of small uncinate hooks present; array 12 hooks wide at base, 14 hooks from base to apex; hooks of array 5-10 (8, n = 5) long; from array, band of larger, uncinate hooks extends distally, disappearing in metabasal region; hooks of band 10-15 (12, n = 5) long.
Segments acraspedote, 1.62-3.70 (2.34, n = 10) mm long, 1.10-1.50 (n = 10) mm wide; length: width ratio 1.18:3.08 (1.86, n = 10); genital pores alternate irregularly, lateral, in posterior third of segment margin, 430-1000 (540, n = 10) (27%) from posterior end. Genital atrium with muscular thickenings anteriorly and posteriorly; hermaphroditic sac pyriform, 300-400 (330, n = 10) long, 130-200 (180, n = 10) wide; cirrus sinuous, unarmed, leading to small, often crescentic internal seminal vesicle, 20-70 (35, n = 10) long, 60-120 (90, n = 10); ovoid external seminal vesicle, thickwalled, 45-120 (79, n = 10) long, 40-80 (58, n = 10) wide, leads to coiled vas deferens. Testes numerous (> 600), 38-53 (44, n = 10) in diameter, filling medulla in single layer; confluent anterior to uterus and posterior to ovaries. Vagina joins cirrus in distal region of hermaphroditic sac, runs along posterior border within hermaphroditic sac; at proximal pole of sac, vagina curves posteriorly towards ovarian isthmus; seminal receptacle absent. Ovary 4-lobed in transverse section, lobes 140-240 (180, n = 10) long, 280-410 (340, n = 10) wide; Mehlis’ gland posterior to ovarian isthmus, 100- 210 (140, n = 10) in diameter. Uterine duct runs anteriorly from Mehlis’ to level of hermaphroditic sac; uterus with thick, glandular walls, extends anteriorly to anterior quarter of segment; anterior extremity of uterus extends ventrally to surface of segment forming potential uterine pore; patency of pore not established. Vitelline follicles circummedullary, obscuring internal details of fully mature segments; follicles 20-30 (24, n = 10) in diameter. Ventral osmoregulatory canals 33-50 (42, n = 5) in diameter; narrow transverse canal, 15 in diameter joins lateral canals at posterior margin of each segment. Dorsal canals not seen in whole mounts or serial sections. Gravid segments absent.
REMARKS
The present redescription, based on new material from New Caledonia, agrees with the original description of Campbell & Beveridge (1993) in most respects. The scolex is shorter in the new specimens and the tentacle sheaths are coiled rather than sinuous, but this is considered to be due to fixation. In the present collection, two poorly preserved specimens were longer than the others and had the tentacle sheaths extended so that they were sinuous rather than coiled. The measurements of these two specimens (not included in the description) were: scolex 8.1, 8.8 mm long; pars bothrialis 0.92, 1.00 mm long; pars vaginalis 6.2, 6.7 mm long; pars bulbosa 1.50, 1.63 mm long. The lengths of these specimens were comparable with the measurements given in the original description, the main difference being in the longer pars vaginalis. The bulb length was not affected by fixation and the lengths of the bulbs of the newly collected specimens were longer than in the original description (1.50-1.82 mm herein; 1.12-1.28 mm in the original description). A prebulbar organ was not present and the original description is considered to be in error. A ring of muscle around the anterior end of the bulb was apparently mistaken for a prebulbar organ. In the original description, the origin of the retractor muscle could not be determined ( Campbell & Beveridge 1993). In the newly collected specimens, the retractor inserts near the anterior extremity of the bulb. The hook sizes in the current redescription are larger than in the original, but this may be due to intraspecific variation. The hooks of paratype specimens were re-examined and the original measurements confirmed. In spite of the differences in measurements, all other features of the tentacular armature were identical with the types.
The principal difference noted in the current description is the presence of a chainette on the external surface of the tentacle. There is one chainette element per principal row of hooks and the chainette elements differ in shape and position from the intercalary row, being aligned with the principal rows rather than posterior to them. On this basis, the species is reallocated to the genus Paragrillotia . Some of these elements were illustrated by Campbell & Beveridge (1993: fig. 12) but were presumably not considered to be significant. The presence of the chainette was confirmed by re-examination of paratype specimens.
In the present description, fully mature segments were available and a uterine pore was detected. In addition, a more detailed description of the terminal genital ducts can be given.
Paragrillotia spratti n. comb. differs from both P. similis and P. apecteta n. sp. in having a single row of intercalary hooks rather than two or three rows, as well as in the basal armature. Paragrillotia apecteta n. sp. has four enlarged hooks in the basal armature which are not present in congeners, while the external surface of the basal armature of P. spratti n. comb. bears a compact array of small hooks not present in P. similis .
The adult of P.spratti n. comb. has previously been reported from the spiral valve of the bronze whaler, Carcharhinus brachyurus (Günther, 1870) , from the coast of New South Wales, Australia ( Campbell & Beveridge 1993) and the plerocercus from the musculature of the teleost Lisa macrolepis (Smith, 1846) from the Arabian Gulf by Kardousha (1999).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Paragrillotia spratti ( Campbell & Beveridge, 1993 )
| Beveridge, Ian & Justine, Jean-Lou 2007 |
Pseudogrillotia spratti
| CAMPBELL R. A. & BEVERIDGE I. 1993: 43 |