
Alpheopsis keijii, Anker, Arthur, 2017
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4226.4.7 |
|
publication LSID |
lsid:zoobank.org:pub:24DA909B-1E1A-4152-B740-5AD96FFB77FE |
|
DOI |
https://doi.org/10.5281/zenodo.6028319 |
|
persistent identifier |
https://treatment.plazi.org/id/03D6774E-FF8F-2B41-FF04-FC86A70E716C |
|
treatment provided by |
GgServerImporter |
|
scientific name |
Alpheopsis keijii |
| status |
sp. nov. |
Alpheopsis keijii View in CoL sp. nov.
( Figs. 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 , 4 View FIGURE 4 a, b)
Alpheopsis trispinosus View in CoL — Banner & Banner 1983: 12. Alpheopsis trispinosus View in CoL — Anker 2001 (vol. 2): 5.
Type material. Holotype: male (cl 4.8 mm), MNHN-IU-2014-12838, Japan, Ryukyu Islands , east of Aguni Island, R / V Toyoshio-Maru sta. 9, 26°30.72’N 127°25.99’E to 26°30.31’N 127°26.13’E, depth: 439–487 m, beam trawl, sand, sponges, 26 May 2002 GoogleMaps . Paratype: ovigerous female (cl 4.5 mm), OUMNH.ZC. 2016-02-024, Japan, eastern Kyushu , off Minami-Hyuga, R / V Toyoshio-Maru sta. 14, 32°20.27’N 131° 48.27E, depth: 136 m, beam trawl, 27 May 2004 GoogleMaps .
Additional material. 1 male (cl 3.3 mm), MNHN-IU-2014-12839, Philippines, Bohol Island, Maribohoc Bay, Panglao Marine Biodiversity Project sta. P1, 9°36.1’N 123°45.0’E, tangle nets, depth: 90–200 m, coll. local fishermen, 30 May 2004 GoogleMaps ; 1 male (cl 3.6 mm), 1 ovig. female (cl 3.7 mm), MNHN-IU-2014-12840, same collection data; 1 male (cl 3.5 mm), OUMNH.ZC. 2016-02-025, Philippines, Bohol Island, Maribohoc Bay, Panglao Marine Biodiversity Project sta. T13, 9°40.5’N 123°49.5’E, depth 90–100 m, sponges, 17 June 2004 GoogleMaps ; 1 ovig. female (cl 4.2 mm), MNNH-IU-2015-426, Papua New Guinea, Solomon Sea, Laughlan Islands, off Budi Budi Island , N.O. “ Alis ” MADEEP campaign sta. DW4287, 09°12’S 153.55°E, depth: 340–375 m, coll. MNHN team, 30 April 2014; 1 male? (pleon damaged, cl 5.0 mm) GoogleMaps , 1 female (cl 4.3 mm), MNNH-IU-2014-18606, Papua New Guinea, Solomon Sea, Laughlan Islands, off Budi Budi Island , N.O. “ Alis ” MADEEP campaign sta. DW4286, 09°12’S 153.55°E, depth: 306–365 m, coll. MNHN team, 30 April 2014 GoogleMaps ; 1 ovig. female (cl 4.3 mm), MNHN-IU-2014- 19481, Mascarene Islands , Réunion, sta. DC 27, no further data, det. Banner as “ A. trispinosus ” (tentative identification, see below).
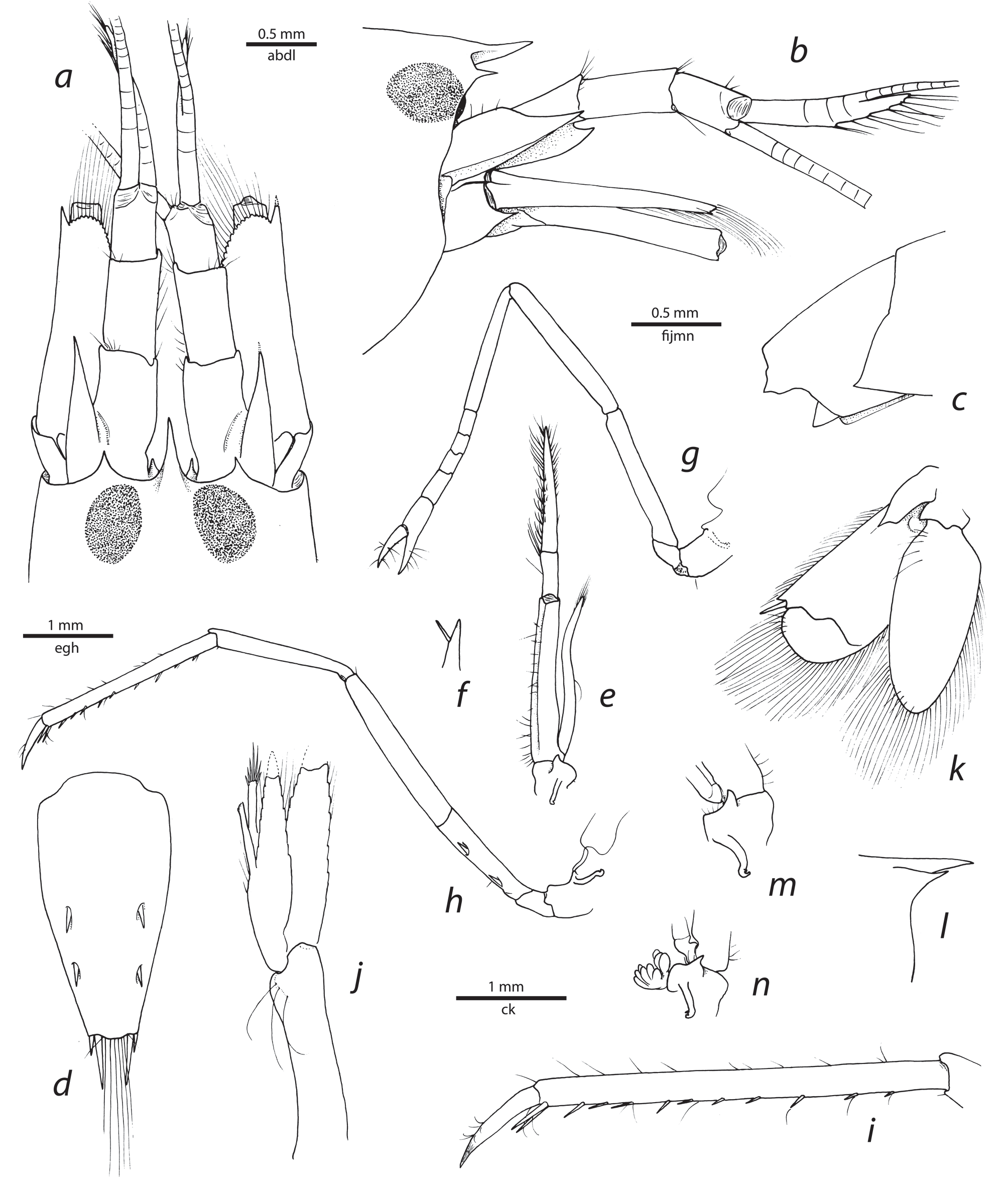
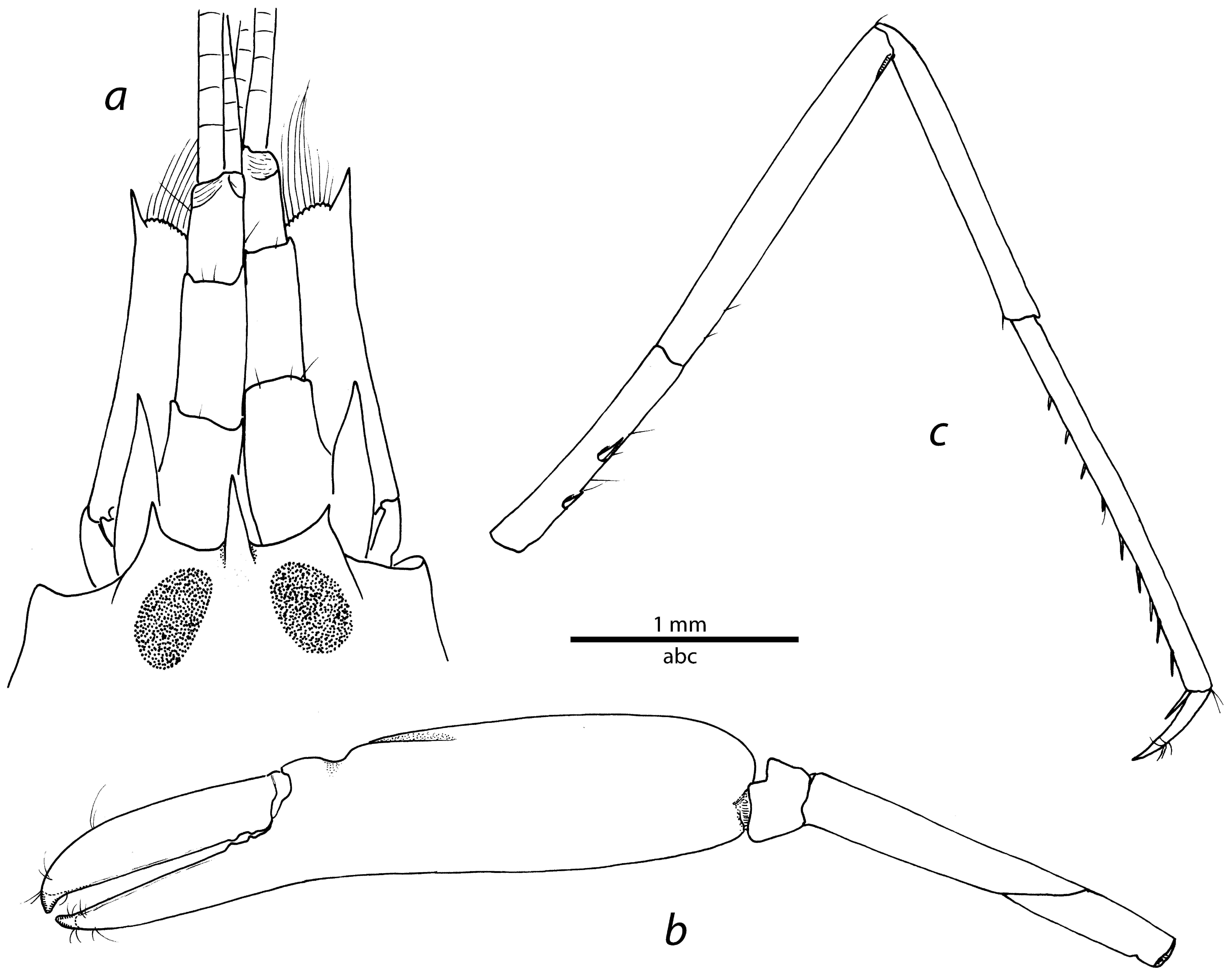
Description. Small-sized species of Alpheopsis (cl <5.0 mm). Carapace glabrous; rostrum much longer than wide, acute distally, base somewhat elevated from adjacent rostro-orbital area, tip not reaching mid-length of first article of antennular peduncle; orbital hoods feebly swollen, with acutely projecting, subtriangular orbital teeth, latter with very wide base; pterygostomial angle rounded, not projecting anteriorly ( Fig. 1 View FIGURE 1 a, b, l).
First to fourth pleura rounded to rounded-angular posteroventrally; fifth pleuron with acutely projecting posteroventral angle; sixth pleonite with subacute posterior lobe and well-delimited, triangular, articulated plate ( Fig. 1 View FIGURE 1 c). Telson relatively narrow, distally tapering, about 2.5 as long as anterior width; dorsal surface with two pairs of spiniform setae inserted at some distance from lateral margin, at about 0.5 and 0.7 telson length, respectively; posterior margin about 0.3 as wide as anterior margin, broadly rounded; each posterolateral angle with one pair of stout spiniform setae, mesial three times as long as lateral ( Fig. 1 View FIGURE 1 d).
Eyes completely concealed by orbital hoods, not visible in dorsal and generally also in lateral view, anteriormost portion occasionally exposed in lateral view; eyestalk with well-pigmented cornea ( Fig. 1 View FIGURE 1 a, b). Epistomial sclerites widely separated, each with small sharp process.
Antennular peduncles relatively slender; stylocerite slender, with acute tip reaching slightly beyond distal margin of first article; ventromesial carina with strong, anteriorly directed, acute tooth; second article about 1.7 times as long as wide; lateral flagellum biramous; fused portion short, with at least three visible joints; accessory ramus long, with more than six groups of long aesthetascs ( Fig. 1 View FIGURE 1 a, b). Antenna with basicerite ending in sharp tooth distoventrally; scaphocerite ovate-elongate, with anteriorly rounded blade and sharp distolateral tooth reaching far beyond anterior margin of blade, but typically typically not exceeding distal margin of antennular peduncle; carpocerite reaching to end of scaphocerite or overreaching it slightly ( Fig. 1 View FIGURE 1 a, b).
Mouthparts typical for genus in external view. Third maxilliped slender, pediform; coxa with distally bilobed lateral plate above mastigobranch, distal lobe more pronounced, subacute; penultimate article about 2.5 times as long as wide; ultimate article tapering to slender corneous tip, with slender spiniform seta subdistally; arthrobranch absent (holotype and paratype) or present (one non-type specimen), in latter case relatively small, with few lamellae ( Fig. 1 View FIGURE 1 e, f, m, n).
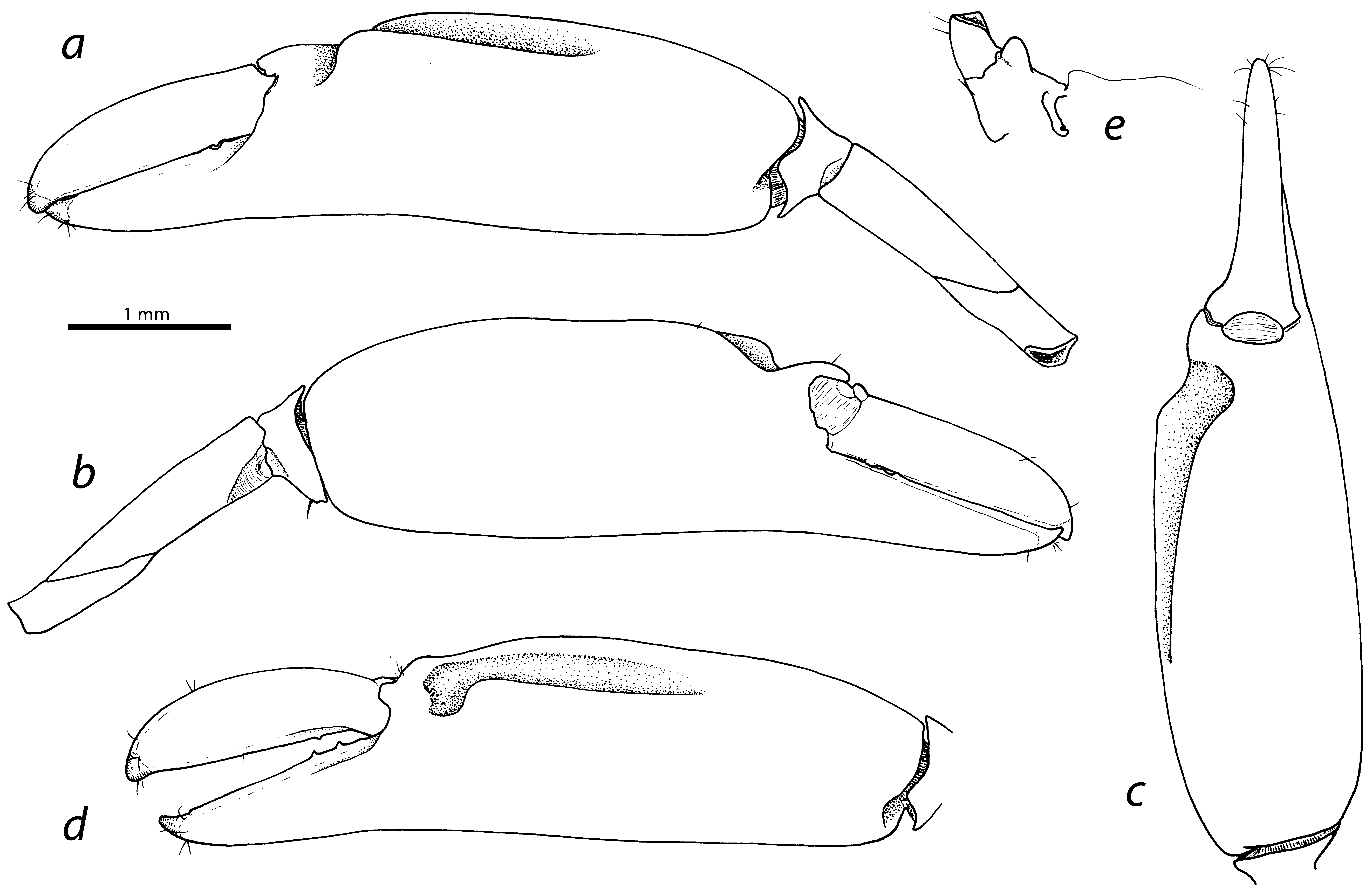
Chelipeds subequal in size, subsymmetrical in shape, carried extended when not in use ( Figs. 2 View FIGURE 2 , 4 View FIGURE 4 b); coxa with rounded lobe dorsally; basis very short; ischium and merus slender, unarmed; merus with dorsal margin about 3.5 times as long as maximum width, distomesial angle blunt; carpus very short, cup-shaped, with rounded distal lobes; chela moderately slender, with fingers about half as long as palm; palm about 2.5 times as long as maximum height, subcylindrical in cross-section, with deep longitudinal groove on dorsal surface, starting at about palm midlength and fusing with short transverse groove close to dactylar articulation; fingers slightly twisted, not gaping when closed, with blunt tips; cutting edge of pollex with two small teeth proximally; cutting edge of dactylus with one small tooth fitting into gap between two teeth of pollex ( Fig. 2 View FIGURE 2 a–e).
Second pereiopod slender; ischium equal to merus in length; carpus with five articles; ratio of carpal articles (from proximal to distal) approximately equal to: 5: 1: 1: 1: 1.5; chela much longer than distal carpal article, with fingers somewhat longer than palm, simple ( Fig. 1 View FIGURE 1 g). Third pereiopod slender; ischium with two short spiniform setae on ventrolateral surface; merus about seven times as long as wide, unarmed; carpus 0.7 length of merus, unarmed; propodus with numerous spaced spiniform setae along ventral margin and two longer spiniform setae distally, adjacent to dactylus; dactylus at most 0.3 length of propodus, slender, simple, subconical, gradually curving distally, with some setae subdistally ( Fig. 1 View FIGURE 1 h, i). Fourth pereiopod generally similar to third pereiopod. Fifth pereiopod missing.
Male second pleopod with appendix masculina reaching far beyond appendix interna, but not reaching tip of endopod; apex with several long stiff setae ( Fig. 1 View FIGURE 1 j). Uropod with lateral lobe of protopod distally bidentate; exopod broadly ovate, with small distolateral tooth; adjacent spiniform seta long, slender; diaresis with subtriangular tooth adjacent to spiniform seta and sinuous mesial portion ( Fig. 1 View FIGURE 1 k).
Gills and exopods as following: five pleurobranchs above first to fifth pereiopods; five mastigobranchs on coxae of third maxilliped and first to fourth pereiopods; five setobranchs on coxae of first to fifth pereiopods; arthrobranch at base of third maxilliped absent or present; podobranch absent; three exopods on first to third maxilliped.
Colour pattern. Background semitransparent whitish; carapace with two broad pale-reddish bands, posterior band broader and crescent-shaped, anterior portion of carapace with reddish markings laterally and dorsally; pleon with six broad, somewhat irregularly shaped, transverse, rusty-reddish bands, one per pleonite, these bands being connected laterally by posterior extension of each band (except on sixth pleonite); chelipeds semitransparent greyish with some orange markings near articulations; walking legs semitransparent; uropod with large rusty-red patch on posterior half; telson with rusty-red markings; ovary and eggs bright yellow ( Fig. 4 View FIGURE 4 a, b).
Type locality. Japan, Ryukyu Islands , east of Aguni Island .
Distribution. Indo-West Pacific: Japan (Kyushu to Ryukyu Islands), Philippines (Bohol Island), Papua New Guinea (Laughlan Islands), Réunion Island (present study), possibly also Madagascar (Nosy-Bé) ( Banner & Banner 1983, as A. trispinosa ) (see below).
Etymology. The new species is named after the author’s colleague and well-known Japanese carcinologist Dr. Keiji Baba, who contributed immensely to the taxonomic knowledge of the decapod fauna of Japan and the world.
Ecology. All specimens of Alpheopsis keijii sp. nov. were collected by trawls or tangle nets at a depth range of 90–487 m. Collecting data for the holotype and one of the non-type specimens from the Philippines indicates that the species may be associated with deep-water sponges.
Remarks. Alpheopsis keijii sp. nov. belongs to the A. trispinosa species group, which also contains A. trispinosa (Stimpson, 1960) from southern and southeastern Australia ( Banner & Banner 1973); A. equidactylus ( Lockington, 1877) from California ( Schmitt 1921; Wicksten 1984); A. chilensis Coutière, 1897 from Chile and Peru ( Retamal 1981; Wicksten & Méndez 1983); A. shearmii ( Alcock & Anderson, 1899) from the Arabian Sea off India ( Alcock & Anderson 1899; Banner & Banner 1977); A africana Holthuis, 1952 from West Africa ( Holthuis 1952); A. diabolus Banner, 1956 from the western Pacific ( Banner 1956; Chace 1988); A. garricki Yaldwyn, 1971 from New Zealand ( Yaldwyn 1971); and A. azorica Anker, d’Udekem d’Acoz & Poddoubtchenko, 2005 from the Azores ( Anker et al. 2005). The A. trispinosa is group is defined mainly by the presence of two deep grooves on the cheliped palm, one longitudinal and one transversal, meeting each other subdistally, and the carapace with acute or (rarely) obtuse orbital teeth ( Coutière 1899; Anker 2001; Anker et al. 2005).
Alpheopsis keijii View in CoL sp. nov. can be separated from both A. trispinosa View in CoL and A. garricki View in CoL also by the absence of strong teeth on the finger cutting edges, on one of the chelipeds (cf. Banner & Banner 1973: fig. 14b; verified on photograph of the holotype of A. garricki View in CoL ); from A. trispinosa View in CoL by the relatively longer second article of the antennular peduncle, being about 1.7 times as long as wide in the new species vs. at most 1.2 times in A. trispinosa View in CoL , and the noticeably more slender dactyli of the third to fifth pereiopods (cf. Banner & Banner 1973: fig. 14a, i); from A. garricki View in CoL by the much shorter stylocerite, barely overreaching the distal margin of the first article of the antennular peduncle (vs. reaching almost to the end of the second article in A. garricki View in CoL ) (cf. Yaldwyn 1971, verified on photograph of the holotype); from A. diabolus View in CoL by the cheliped dactylus as long as pollex, and not particularly curved or twisted (vs. much longer than pollex and distally strongly curved and somewhat twisted in A. diabolus View in CoL ) and the simple dactyli of the third to fifth pereiopods (vs. strongly biunguiculate in A. diabolus View in CoL ) (cf. Banner 1956: fig. 3c–e, h; in addition to the personally examined material from Kiribati); from A. shearmii View in CoL by the not particularly swollen orbital hoods, a noticeably weaker armature on the cheliped fingers and a broader telson, the latter also with a very slightly convex posterior margin (vs. with the conspicuously swollen orbital hoods, a stronger armature on the cheliped fingers, and the telson more slender and with a strongly convex posterior margin, in A. shearmii View in CoL ) (cf. Banner & Banner 1977: fig. 1a, c–e); from both A. africana View in CoL and A. azorica View in CoL by the subequal chelipeds (unequal in the other two species, cf. Anker et al. 2005: fig. 1a); from A. africana View in CoL by the noticeably shorter stylocerite, barely overreaching the distal margin of the first article of the antennular peduncle (vs. reaching distinctly beyond this margin in A. africana View in CoL ), and the longer scaphocerite, with its blade reaching well beyond the distal margin of the second article of the antennular peduncle (vs. barely overreaching it in A. africana View in CoL ) (cf. Holthuis 1952: fig. 11a; Anker et al. 2005: fig. 5a, b); from A. azorica View in CoL by the shorter orbital hoods and rostrum, the latter not reaching midlength of the first article of the antennular peduncle (vs. reaching far beyond the distal margin of the first article in A. azorica View in CoL ), the much more slender antennular peduncles, including a comparatively longer second article (cf. Anker et al. 2005: fig. 1b), and the presence of strap-like epipods (mastigobranchs) on the coxae of the first to fourth pereiopods (vs. on the coxae of the first and second pereiopods in A. azorica View in CoL ); from A. chilensis View in CoL by the distally sharp orbital teeth (vs. obtuse in adult specimens of A. chilensis View in CoL ; in very young specimens of A. chilensis View in CoL (cl <5 mm), the orbital teeth are distinctively pointed) (cf. Coutière 1899: fig. 27; Wicksten & Méndez 1983: fig. 3; present study); and from both A. chilensis View in CoL and A. equidactylus View in CoL by the finger cutting edges of the chelipeds without strong teeth (vs. with strong teeth in the other two species), the proportionally much longer second article of the antennular peduncle, being about 1.7 times as long as wide (vs. as long as wide in A. chilensis View in CoL or 1.2–1.3 times as long as wide in A. equidactylus View in CoL ), and the much shorter stylocerite, barely overreaching the distal margin of the first article of the antennular peduncle (vs. reaching almost to the distal margin of the second article in A. chilensis View in CoL and at least to the mid-length of the second article in A. equidactylus View in CoL ) (cf. Wicksten & Méndez 1983: figs. 3, 5; Wicksten 1984; present study).
Hale (1941) provided illustrations of an ovigerous female (cl 7.0 mm, total length: 17 mm) dredged at 122 m off eastern Tasmania that he tentatively identified as “ Alpheopsis trispinosus View in CoL ”, stating that although it “may be separable from A. trispinosus View in CoL , one hesitates to propose a new name because of the paucity of the material”. Hale also compared his Tasmanian female with the description of an incomplete juvenile specimen (total length: 8.2 mm) from Indonesia by De Man (1922), concluding that the two specimens “agree quite closely”. However, they differ greatly in the proportions of the antennular peduncles and therefore are extremely unlikely to be conspecific. The identity of the Tasmanian specimen remains unknown; it may represent represent A. garricki View in CoL , a morphologically slightly different deep-water form of A. trispinosa View in CoL , or an undescribed taxon. In any case, A. keijii View in CoL sp. nov. differs from A. trispinosa sensu Hale (1941) View in CoL by the much shorter stylocerite (cf. Fig. 1 View FIGURE 1 a and Hale 1941: fig. 4A).
When alive, A. keijii sp. nov. may be distinguished from A. trispinosa , A. garricki , A. chilensis and A. equidactylus by some details in the colour pattern ( Fig. 4 View FIGURE 4 a, b), e.g. by the transverse bands being laterally connected (which is not the case in the other four species, examples in Fig. 4 View FIGURE 4 c–f); the colouration of the remaining species of the A. trispinosa complex remains unknown. Based on the above comparison, A. keijii sp. nov. appears to be closest to the eastern Atlantic A. africana and a genetic comparison of the two taxa is desirable to confirm the validity of the new species.
Alpheopsis keijii View in CoL sp. nov. appears to be variable in the development of an arthrobranch at the base of the third maxilliped: it is absent in the holotype and paratype from Japan and present, albeit feebly developed, in at least one specimen from the Philippines ( Fig. 1 View FIGURE 1 g, n). The presence or absence of an arthrobranch is generally constant within most alpheid genera. However, as noted by Banner & Banner (1973) and Miya (1981), some species of Alpheopsis View in CoL do have an arthrobranch, while others do not, and therefore, an intraspecific variation of this character would not be very surprising. However, careful dissection of additional Japanese specimens of A. keijii View in CoL sp. nov. is needed to confirm that the arthrobranch is indeed absent and was not accidentally detached while dissecting the two type specimens. Furthermore, DNA analyses are needed to confirm the conspecificity of the Japanese and the Philippine specimens and to reject the alternative hypothesis that two closely related species are involved, one with and one without arthrobranch.
The ovigerous female from Réunion ( Fig. 3 View FIGURE 3 ) presents almost all diagnostic features of A. keijii View in CoL sp. nov., except for the noticeably more slender chelipeds (compare Figs. 2 View FIGURE 2 a, b, 3b), which could be explained by sexual dimorphism or other form of intraspecific variation, perhaps depth-related. This specimen is therefore tentatively included under additional material of the new species, awaiting confirmation. Banner & Banner (1983) reported A. trispinosa View in CoL from Madagascar based on a single specimen dredged at a depth of 205 m off Nosy-Bé. Although this specimen could not be located in the MNHN collection, its geographical and bathymetric collection data strongly suggests that it is not A. trispinosa View in CoL , which appears to be restricted to the temperate and shallower waters (11–135 m) of New South Wales, Victoria, Tasmania, South and Western Australia ( Hale 1941; Banner & Banner 1973, 1982; Poore 2004), but instead may well belong to the herein described new species. The presence of A. keijii View in CoL sp. nov. in the southwestern Indian Ocean will need to be confirmed by additional sampling and photography of deepwater alpheids in this biogeographically interesting area. As already noted by Anker et al. (2005), the record of A. trispinosa View in CoL from Florida by Gore (1981) also refers to a different species, probably undescribed and more closely related to A. africana View in CoL and A. azorica View in CoL than to A. trispinosa View in CoL ; this species is included in the key below as A. sp. aff. trispinosa View in CoL (see below).
The validity of A. garricki from New Zealand was questioned by Yaldwyn (as pers. comm. to Banner & Banner 1973), who believed that it may well be a junior synonym of A. trispinosa . The type material of A. garricki was collected in the Cook Strait and off Chatham Islands, at a depth range of 36–200 m, which is similar to that of A. trispinosa (see above). The colour patterns of A. trispinosa ( Fig. 3 View FIGURE 3 c) and A. garricki (described in Yaldwyn 1971) appear to be similar if not identical. Based on Yaldwyn’s (1971) very brief description and a photograph of the holotype of A. garricki supplied by the NMNZ, the two nominal species appear to differ only in the length of the stylocerite, which reaches almost to the distal margin of the second article of the antennular peduncle in A. garricki vs. at most to the mid-length of the second article in the neotype of A. trispinosa (cf. Banner & Banner 1973: fig. 14a). Furthermore, Banner & Banner (1973) reported some variation in their Australian material of A. trispinosa , e.g. in the shape of the frontal margin of the carapace, illustrating the narrower and distally more acute orbital teeth in a 16 mm female dredged from about 135 m off New South Wales. Therefore, it cannot be excluded that Banner & Banner’s (1973) material contained two species, A. trispinosa and a second species, perhaps A. garricki . For these reasons, it seems best to treat A. garricki as a valid species, until more evidence is available.
The preliminary key below provides an identification aid to the nine currently known species of the Alpheopsis trispinosa complex and also includes A. trispinosa sensu Gore (1981) as a separate, possibly undescribed taxon, A. sp. aff. trispinosa . The number of strap-like epipods on the pereiopods remains unknown in at least two species of the A. trispinosa group and therefore could not be included in this key; however, the reduced number of epipods represents an additional diagnostic character of A. azorica ( Anker et al. 2005; see below). The separation of A. equidactylus from A. garricki on the basis of the slightly different length-width ratio of the third pereiopod propodus is highly tentative and needs confirmation. In fact, both taxa need full redescriptions (see above); however, since they are geographically widely separated, they cannot be confused provided correct locality data is available.

| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Alpheopsis keijii
| Anker, Arthur 2017 |
Alpheopsis trispinosus
| Banner 1983: 12 |