
Chironius challenger, Kok, Philippe J. R., 2010
|
publication ID |
https://doi.org/ 10.5281/zenodo.197801 |
|
DOI |
https://doi.org/10.5281/zenodo.5630130 |
|
persistent identifier |
https://treatment.plazi.org/id/03D71055-C948-6C01-FF66-34DDFA88FD14 |
|
treatment provided by |
Plazi |
|
scientific name |
Chironius challenger |
| status |
sp. nov. |
Chironius challenger sp. nov.
( Figs. 1 View FIGURE 1 , 2 View FIGURE 2 A–C, 3 A–D, 4 D, 6, 7; Tables 1 View TABLE 1 , 2 View TABLE 2 )
Chironius fuscus (Linnaeus, 1758) ; Mägdefrau et al. (1991: 22 –24, figs. 22a, b) Chironius fuscus fuscus (Linnaeus, 1758) in part; Dixon et al. (1993: 114)
Chironius fuscus (Linnaeus, 1758) ; Myers & Donnelly (2008: 121, fig. 73)
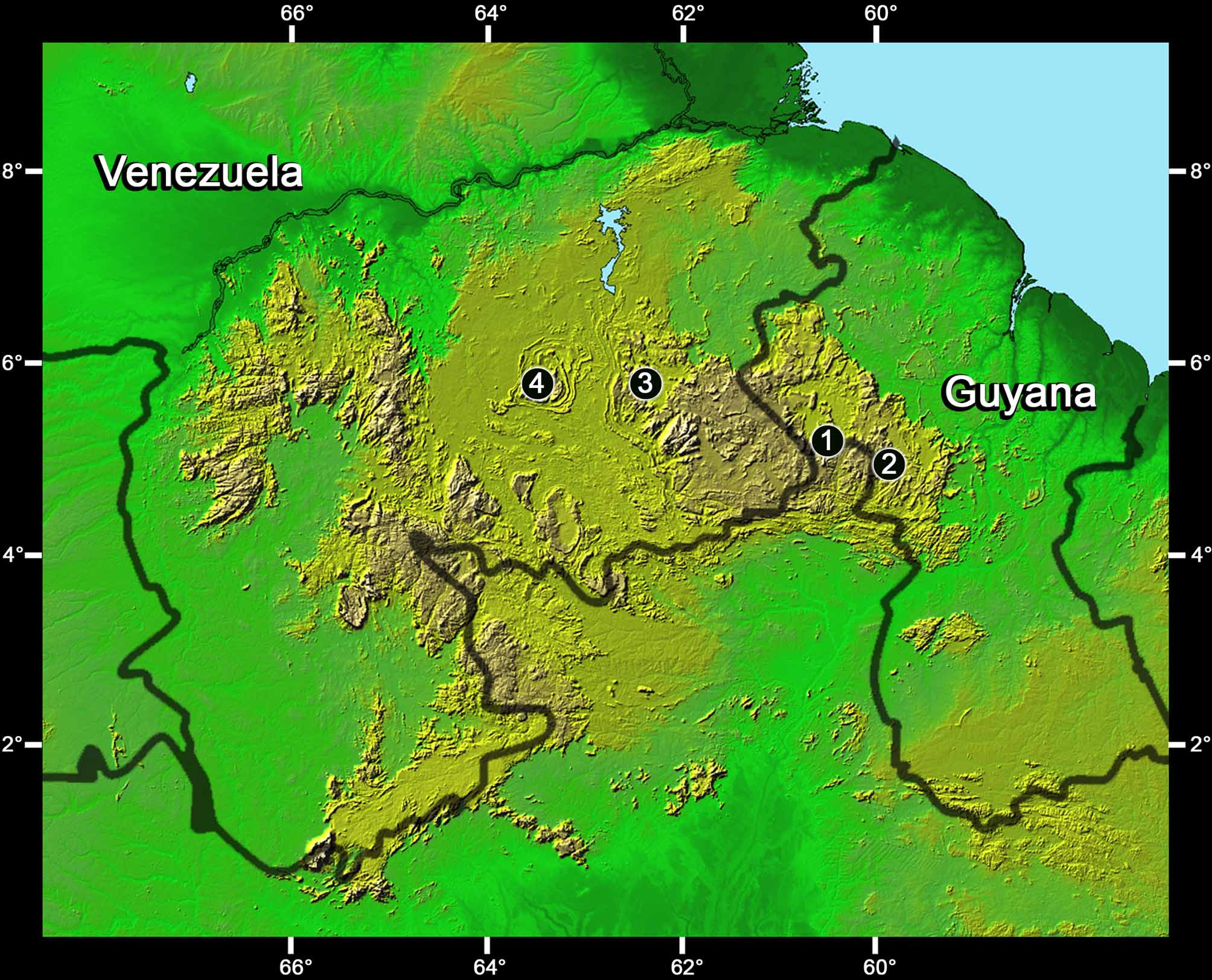
Holotype. IRSNB 2659 (field number PK 2017), a subadult female collected by Philippe J. R. Kok, 25 November 2007 at 10h45, southeastern slope of Maringma Tepui, Cuyuni-Mazaruni District, Guyana (05° 12’ N, 060° 35’ W, 1500 m elevation).
Referred specimens. BMNH 1976.235, adult female collected by Adrian Warren, 17 August 1974, southern side of Auyantepui, 2088 m elevation, Venezuela; ROM 42603, juvenile collected by Samir Khan, 2 November 2004, Mount Wokomung, 1400 m elevation, Guyana; MHNLS 16088, subadult female collected by Gilson Rivas, 8 February 2002, Auyantepui, 1700 m elevation, Venezuela.
Etymology. The specific epithet is considered to be a noun in apposition and refers to Professor George Edward Challenger , the fictional main character of Sir Arthur Conan Doyle's famous novel "The Lost World" published in 1912. "The Lost World" depicts an epic expedition to an isolated tepui in the middle of the tropical jungle of northern South America in search of dinosaur-like creatures and a forgotten civilisation. The reading of Sir Conan Doyle’s novel when I was a child fuelled my thirst for exploration and adventure and positively influenced my research and interest in the Pantepui region.
Definition and diagnosis. A species of the genus Chironius characterized by the following combination of characters: 10 dorsal scale rows at midbody, absence of apical pits and paravertebral keels in female (male unknown), 152–159 ventrals, 99–103 subcaudals, anal plate single, loreal as long as—or slightly longer than—high, 39–41 maxillary teeth.
Only five of the 20 currently known Chironius species occur in the Guiana Shield (area sensu Ávila-Pires 2005, number of species f i de Hollis 2006): C. carinatus (Linnaeus, 1758) , C. cochranae Hoge and Romano, 1969 , C. exoletus (Linnaeus, 1758) , C. fuscus (Linnaeus, 1758) , and C. scurrulus (Wagler, 1824) . Among these species, only C. fuscus and C. scurrulus have 10 DSR at midbody, a condition shared by C. challenger . All other species have 12 DSR at midbody.
Chironius challenger is readily distinguished from C. scurrulus by the following characters (characters of C. scurrulus in parentheses): absence of apical pits (present, at least on neck), loreal as high as long or only slightly longer than high (distinctly longer than high), juveniles brownish with lighter crossbands (uniform green in life, blue-black in preservative), adults brownish with lighter crossbands (red or reddish brown in life, colouration variable in preservative, but never with lighter crossbands).
Chironius challenger is mostly distinguished from C. fuscus by the following characters (characters of C. fuscus in parentheses): absence of apical pits (present, at least on neck), absence of paravertebral keels (usually present, even if faint in females), loreal as high as long or only slightly longer than high (distinctly longer than high), higher number of ventrals - minimum 152 in C. challenger (maximum 149 in C. fuscus from the Guiana Shield), shorter tail - 30–31% TTL in C. challenger (34–37% TTL in C. fuscus from the Guiana Shield), lower number of subcaudals - maximum 103 in C. challenger (minimum 118 in C. fuscus from the Guiana Shield), infralabials 9 (usually 10), ventrals in adults conspicuously mottled with darker edges (not heavily mottled, no conspicuous darker edge), skin between scales blue in life (white). Further comparisons are given in the Discussion.
Description of the holotype. A subadult female (according to oviduct development), in good condition, TTL 720 mm, SVL 500 mm, TL 220 mm (30.6% TTL); head well distinct from neck, narrow anteriorly, widest at temporal region, HW 54.8% of HL; snout rounded in dorsal and lateral view, SL 6.2 mm; eye large (EL 4.8 mm), pupil round, EN 19.3% HL; rostral large, broader than long (width about 1.8 times height), visible from above; internasals squarish, as broad as long, slightly smaller than prefrontals, laterally in contact with nasal, their medial suture asymmetrical, noticeably sinistral to the prefrontal medial suture; prefrontals slightly larger than internasals, width 1.5 times length, irregularly hexagonal, separated from eye by preocular and supraocular; frontal pentagonal, bell-shaped, longer than wide, 3.0 times length of medial prefrontal suture; parietals large, length 1.5 times maximal width, medial suture of parietals and length of frontal equal; 5 postcephalic scales. Nasal scale partially divided dorsally and ventrally, posterior part as long as, but slightly higher than, anterior part; preocular 1/1, separated from frontal by prefrontal-subocular contact; loreal slightly longer than high (height 93% length), in contact with nasal, prefrontal, preocular, second and third supralabials, separated from orbit by preocular; suboculars absent; postocular 1/1; temporals 2+2/1+1, supralabials 9/9, sixth largest, first contacting nasal, second contacting nasal and loreal, third contacting loreal and in point contact with preocular, fourth, fifth, and sixth entering orbit, sixth in point contact with postocular and contacting anterior temporal, seventh and eighth contacting anterior temporal, ninth contacting anterior and posterior temporals; infralabials 9/9, first seven in contact with genials, first pair meeting at ventral midline, separating mental from genials; 2 pairs of genials, the first shorter. Maxillary teeth 39. Dorsal scales in 10-10-10 rows, smooth without apical pits, a few weak anal ridges laterally above cloacal region; ventrals 159; caudal scale row reduction from 6 to 4 rows occurs at the 5th subcaudal; anal single; divided subcaudals 103 ( Figs. 1 View FIGURE 1 , 2 View FIGURE 2 A–C, 3 A, 4 D).
Colour of the holotype in life. Dorsolateral body ground colour light brown, darkening posteriorly, with a series of ca. 31 ill-defined greyish brown transverse bands (1, rarely 2 scales wide) outlined in black. Tail darker brown without conspicuous transverse bands. Dorsal scales with black edges, sometimes slightly peppered with dark brown or black; some dorsal scales yellow anteriorly, especially on anterior body and on lower flank. Skin between scales sky blue. Head light brown, supralabials yellow, with labials 2–3 having dark posterior edges; black postocular stripe from eye across lower part of primary temporal and upper part of ultimate supralabial reaching postcephalic scales then breaking into blotches on the lateral side of ventrals; iris dark copper; tongue black. Underside of head immaculate yellowish white; ventrals immaculate yellow until ca. the 36th, then posteriorly bordered by black, mottled with brown and black spots increasing in number posteriorly, becoming almost completely black near the vent; subcaudals similar in colour and pattern, but less heavily mottled by black than the posteriormost ventral scales ( Figs. 1 View FIGURE 1 A–B, 4 D).
Colour of the holotype in preservative. Pattern essentially as in life, but dorsal colours faded to grey and light grey, and yellow and sky blue faded to white ( Figs. 1 View FIGURE 1 C–D, 2 A–C, 3 A, 6).
Variation. Three referred specimens are available, consisting of two females and a juvenile. The females vary in TTL from 720–937 mm (315 mm in the juvenile), in SVL from 450–658 mm (222 mm in the juvenile), and in TL from 201–279 mm (93 mm in the juvenile). Tail length is 42–45% SVL. The loreal is always high, its height being 93 (in the holotype)–100% (in all other specimens) its length. There is little variation in scale counts. Weak anal ridges laterally above the cloacal region are absent in one female (MHNLS 16088) and the juvenile (ROM 42603). Ventrals range from 152–159, divided subcaudals 99–103 (65% number of ventrals), supralabials 8–9, number of infralabials in contact with genials 5–7, postcephalics 4–6. Although the holotype has only 1 postocular on both sides, all other individuals have 2. The temporal formula is quite variable and only one specimen (BMNH 1976.235) has 1+1 temporal on both sides. Other specimens have 1+1/1+2 (MHNLS 16088), 1+2/1+2 (ROM 42603) or 2+2/1+1 (holotype) temporals. Paravertebral scales are smooth and apical pits are absent in all specimens. Caudal scale row reduction from 6 to 4 rows occurs at the 5–7th subcaudals in females, at the 10th subcaudal in the juvenile. Maxillary teeth are 39–41 in females, 41 in the juvenile. See Table 1 View TABLE 1 for measurements and scalation for the holotype and the referred specimens.
Colour and pattern are similar between the referred specimens and the holotype. Intensity of ventral mottling seems related to age and size, the two largest individuals having the most mottling on ventrals. Transverse bands are much more conspicuous and more numerous (ca. 55) in the juvenile specimen. Larger specimens mostly lack transverse bands on the tail whereas these bands are conspicuous in smaller individuals.
Distribution and ecology. Based on museum specimens, Chironius challenger is known between 1400– 2088 m elevation from three localities in the eastern Pantepui region: Auyantepui in Venezuela, Maringma Tepui and Mount Wokomung in Guyana ( Fig. 5 View FIGURE 5 ). An additional specimen collected at 1520 m elevation on the summit of Guaiquinima Tepui, Venezuela ( Fig. 5 View FIGURE 5 ) is illustrated in Mägdefrau et al. (1991: 22, 23) under C. fuscus (see Discussion and additional comparisons).
The holotype was collected in the morning while it was moving in a small tree about 200 cm above the ground in low evergreen upper-montane forest. ROM 42603 was collected while moving on the ground on the northeastern slope of Mount Wokomung, also in low evergreen upper-montane forest. No data on microhabitat is available for the two other referred specimens, which are presumably from low evergreen upper-montane or high-tepui forest according to the collection localities. The specimen reported by Mägdefrau et al. (1991) from Guaiquinima Tepui was collected in the afternoon, on a rock on the bank of a stream dissecting a rocky area covered by Stegolepis (Rapateaceae) and small patches of forest (presumably high-tepui forest given the elevation).
The holotype and MHNLS 16088 have few enlarged ovarian follicles (largest 3.3 and 1.9 mm in length, respectively), and BMNH 1976.235 has three eggs (11.6, 13.6, and 17.1 mm in length).
Sex Female Female Female Juvenile
Locality Maringma Tepui, Auyantepui, Auyantepui, Mt. Wokomung,
Guyana Venezuela Venezuela Guyana


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.