
Halecium flexile Allman, 1888
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3852.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:7DE3BCBA-E5F0-4F0D-B2FD-B5B59E4DAE51 |
|
DOI |
https://doi.org/10.5281/zenodo.6143171 |
|
persistent identifier |
https://treatment.plazi.org/id/03D92A2C-4763-FFCA-FF51-86B85AA1A015 |
|
treatment provided by |
Plazi |
|
scientific name |
Halecium flexile Allman, 1888 |
| status |
|
Halecium flexile Allman, 1888 View in CoL
(Plate 2A; Figs 2A, B View FIGURE 2. A, B ; 3A; 4A; Table 4 View TABLE 4 )
Halecium flexile Allman, 1888: 11 View in CoL , pl. 5 Fig. 2 View FIGURE 2. A, B .― Ralph, 1958: 337, Fig. 12L– O. Halecium delicatulum View in CoL ― Millard, 1977: 7, Fig. 1 View FIGURE 1. A, B C, D.―Galea, 2007 (pro parte): 54, Fig. 12I, J (not Fig. 12F–H = H. platythecum View in CoL sp. nov.).
not Halecium delicatulum Coughtrey, 1876a: 299 View in CoL ; 1876b: 26, pl. 3 Figs 4 View FIGURE 4. A ‒ F, M , 5 View FIGURE 5. A ‒ E .
Material examined. HF9, Isla Santi Bañez, lat. -54.91725, long. -68.36317, 29.5 m, 16.xii.2010, sample C252: a 12 cm high sterile colony (MHNG-INVE-86247). HF9, NE of Channel Murray, lat. -54.95097, long. -68.40114, 16.xii.2010, 18 m, sample C257: two sterile colonies, 9 and 13.5 cm high, respectively (MHNG-INVE-86249). HF11, SW of Isla Nalcayec, lat. -46.10075, long. -73.88970, 29.xi.2011, 26.5 m, sample C317: numerous polysiphonic stems, up to 7 cm high on dead gorgonian, some bearing either male or female gonothecae (MHNG- INVE-86248). HF11, Punta Garro, lat. -46.31517, long. -75.65203, 30.xi.2011, 17.3 m, sample C349: several polysiphonic stems, up to 7.5 cm high, some bearing either male or female gonothecae (MHNG-INVE-86250). Punta Arenas, Faro San Isidro, lat. -53.78174, long. -70.97391, 25.ii.2010, 10 m, sample PTA002: several polysiphonic stems and fragments, up to 8.5 cm high, some bearing male gonothecae (MHNG-INVE-86252). New Zealand, Wellington, Te Raekaihau, lat. -41.3475, long. 174.7897, 1994, 0–2 m, female colony of Halecium delicatulum Coughtrey, 1876 mounted on microslide6 (MHNG-INVE-26669).
Description. Colonies arising from creeping, tortuous, branched, anastomosing stolonal fibers grouped together so as to form a fascicled hydrorhiza. Young stems monosiphonic, mostly found at the periphery of colonies, being rapidly invested by additional tubes, thus giving rise to large, branched, polysiphonic colonies (Pl. 2A). Stems divided into internodes by oblique nodes slanting in opposite directions. Internodes almost collinear ( Fig. 2A View FIGURE 2. A, B ), even in the youngest stems, moderately long, with 2–3 basal annuli and a distal bulge; each giving rise to a well-developed latero-distal hydrophore, reaching the level of the distal node; a pseudodiaphragm on adaxial side ( Fig. 2B View FIGURE 2. A, B ). Side branches arise singly from below the primary hydrophores; alternate, generally given off in the same plane or slightly displaced towards the front and/or rear side of the stem, giving the colony an almost planar appearance. Up to third-order branches in a well-developed colony. Hydrothecae borne on corresponding hydrophores, generally renovated up to four times; rather deep, margin strongly everted. Gonothecae given off from the primary hydrophores. Male ovoid, tapering in the lower third into indistinct pedicel, laterally flattened, with no evident aperture (Fig. 3A4). Female pyriform and greatly swollen, tapering basally into indistinct pedicel; distally, a centrally-placed, rounded aperture (Fig. 3A1–3). Cnidome ( Fig. 4A View FIGURE 4. A ‒ F, M ): microbasic heteronemes [(5.2–5.4) × 2.6 µm] and microbasic mastigophores [(6.6–6.9) × 2.0 µm]
PLATE 2. Preserved colony of Halecium flexile Allman, 1888 with numerous stems growing on axis of dead gorgonian (A), H. maximum sp. nov. (B), H. modestum sp. nov. (C), H. platythecum sp. nov. (D), and H. tehuelchum (d’Orbigny, 1842) (E). All photos by H.R.G. Scale bars: 2 cm (A–C) and 1 cm (D, E).
Remarks. The material on which Allman (1888) established his species was gathered from two remote localities, viz. Marion Island and Tierra del Fuego. Totton (1930, p. 145) selected as the holotype the fertile specimen originating from the former locality. According to him, the Patagonian material “is similar to it but the hydranths are larger and the distal part of the hydrophore is deeper, measuring 0.075 mm compared with 0.050 mm; and it lacks gonothecae”. This statement, of poor informative value, casts some doubt on the identity of both specimens, instead of clarifying their relationships.
Though rather concise, the account by Millard (1977) provides interesting insights into the knowledge of the present species. Her material from Crozet7, assigned to H. delicatulum Coughtrey, 1876 , possesses female gonothecae that are indistinguishable morphologically from those occurring in the Chilean samples, and contain a similar number of large oocytes. In addition, its occurrence on gorgonians (a common substrate for the Chilean specimens) suggests that our material most probably belongs to the same species.
Taken together, the accounts by Allman and Millard, the illustrations of the type material by Ralph (1958, Fig. 12L– O), as well as the present observations, allow us to provide the following diagnosis for H. flexile : colonies with polysiphonic stems, pinnately branched in almost one plane, composed of moderately long, collinear internodes, each bearing a well-developed hydrophore provided with an adaxial pseudodiaphragm and topped by a hydrotheca with everted rim; female gonotheca pyriform, conspicuously swollen distally, male ovoid and flattened laterally. According to that, it will be shown below that the colonies with monosiphonic stems examined earlier by Galea (2007, as H. delicatulum ) belong to a distinct species, H. platythecum sp. nov.
The taxonomic position of several closely-related species of Halecium from the southern hemisphere has been a complex matter of debate for a long time, and the opinions of the taxonomists that have examined the type of H. flexile contrast radically. On one hand, Billard (1910) reached the conclusion that the nominal species H. gracile Bale, 1888 is not more than a synonym of it, with the former having priority. On the other hand, Totton (1930) rejected their synonymy, arguing that “ H. gracile is of smaller proportions, and the male gonothecae are considerably smaller and of different shape”.
Meanwhile, Bale (1915, p. 246) followed Billard and included in the synonymy of H. flexile not only his H. gracile , but also his H. parvulum Bale, 1888 , the latter being considered as the full growth form of H. gracile . Ultimately, other authors [e.g. Ralph (1958), Millard (1975), Watson (1997, 2003a, 2008)] broadened the synonymy of this species group, in considering H. delicatulum as the oldest available name, and sunk in its synonymy the three nominal species mentioned above.
Following examination of various samples from Chile, New Zealand, and southern Australia, it becomes clear that several morphologically similar species were lumped together as H. delicatulum .
First of all, according to Ralph (1958) and the observations made by one of us (H.R.G.) on New Zealand material (sample MHNG-INVE-26669), the following diagnosis for H. delicatulum given by Ralph (1958, p. 337) should be followed scrupulously: “short (up to 3.0 cm) erect monosiphonic stems with loose irregular branching; […] nodes twisted obliquely and sloping alternately in opposite directions; hydrothecal margin strongly everted; […] mature male gonotheca elongate oval, about 0.75 mm in length; mature female gonotheca about 1.0 mm in length, more or less oval in general outline, with two distinct peaks arising from the narrow sides at the distal end; aperture, in depression between the peaks”. Since all the New Zealand material studied by Ralph had colonies with monosiphonic stems, it is worth stressing that this is most probably the normal habit of this species. Accurate, recent illustrations of H. delicatulum were given by Schuchert (2005) and Galea & Schories (2012b). Consequently, only those specimens of Halecium whose mature female gonothecae are laterally flattened and have the aperture situated “between two distinct blunt, heavy, tooth-like processes” ( Ralph 1958, p. 336) should be considered as belonging to Coughtrey’s species. The drawing in Fig. 3B allows the shape and size of the female gonotheca to be compared to those of related species. An additional record of H. delicatulum is, most probably, that of Marktanner-Turneretscher (1890, as H. parvulum 8), but possibly not that of Hartlaub (1901a9) (this assumption
7. The Crozet shelf, situated about 1000 km away from Marion Island, lets us to assume that the specimens examined by Millard (1971) were possibly conspecific with H. flexile .
8. The synonymy is mainly based on the typical shape of the female gonotheca and its high number and small size of oocytes depicted by Marktanner-Turneretscher (1890) in his Pl. 3 Fig. 22.
9. The trophosome in Hartlaub’s (1901a) material was poorly illustrated, and the characteristic pseudodiaphragm was not depicted. In addition, the presence of a male colony is of poor informative value, and should be interpreted with caution. In addition, Ralph’s (1958) reading of Hartlaub’s text is obviously a misinterpretation, adding information that is missing from his account. Hartlaub clearly stated the following: “Das Material […] besteht nur aus einem kleinen Stück Rhizom, von welchem eine Gonothek entspringt”, which means that only one gonotheca, arising from the stolon, was present in the material studied. Although the gonotheca was originally described as broadly rectangular and laterally flattened (thus possibly male), Ralph stated that Hartlaub would have seen some gonothecae “rectangular in general outline and others with two peaks at the distal end”, thus concluding that “some at least of Hartlaub’s specimens of H. delicatulum were female in sex”.
considers the trophosome alone), though both are from New Zealand. Based on the data from the literature, it is therefore assumed that the species may be endemic to New Zealand.
Second, the haleciids with polysiphonic stems reaching up to 10 cm in height, included by Ralph in her diagnosis of H. delicatulum (p. 336), are characters belonging to other nominal species (viz. H. flexile and H. parvulum ) that were sunk in its synonymy. Besides the lack of information on the female gonotheca of Allman’s species, Ralph concluded that the “complete similarity of internode, node, hydrotheca and male gonotheca of H. delicatulum and H. flexile leaves little doubt that the latter is a synonym of H. delicatulum ”. However, due to the lack of suitable material, she failed to explain “what factors cause the readily distinguishable differences in growth form and size”, the nature of these factors being not invoked (e.g. ecological, geographical etc.).
Third, the southeastern Australian material mentioned by Galea (2010b, p. 8) was re-examined for this study. The colonies are exclusively composed of monosiphonic stems up to 3.5 cm high, their branching is irregular and attains the third order, the internodes are slightly zigzagging ( Fig. 2 View FIGURE 2. A, B C), the primary hydrophores are rather short, but still exhibit a conspicuous pseudodiaphragm on their adaxial side, and the hydrothecal rim is only slightly everted ( Fig. 2 View FIGURE 2. A, B D). Terminal stolonization occur quite abundantly, giving rise to additional erect stems, and the gonothecae are given off from both the stolons and stems. The cnidome ( Fig. 4B View FIGURE 4. A ‒ F, M ) is composed of three capsules: small (ca. 3.4 × 1.4 µm), seed-shaped and large [(8.2–9.0) × 2.6 µm], banana-shaped macrobasic heteronemes, as well as a microbasic mastigophore (ca. 6.5 × 2.0 µm). A closer inspection of the gonothecae, previously described as female, revealed that they are, in fact, male10. They are broadly ovoid in frontal view (Fig. 3C) and lenticular when seen from above. Those that are already spent exhibit a small, apical, circular aperture resulting from the local perisarc being ruptured upon the release of gametes (Fig. 3C2). Since no female colonies could be observed, we assign, with a query, the present material to H. balei 11 Fraser (1911). If our specimens indeed belong to it, then Totton’s (1930) argument (see above) supporting its separation from H. flexile is entirely justified [compare Figs 2A and 2 View FIGURE 2. A, B C, as well as Fig. 3A4 and Bale’s (1888) Fig. 3]. Moreover, the relationships between H. balei and H. parvulum should be clarified in light of modern taxonomy.
Fourth, the relationships between H. antarcticum Vanhöffen, 1910 and H. flexile have been questioned, among others, by Totton (1930) and Galea & Schories (2012b). According to Peña Cantero (2014), the female gonothecae of the former are comparatively larger, flattened fronto-dorsally, tapering rather abruptly below, with a wide distal aperture, leaving the passage of a large acrocyst, thus considerably different from Allman’s species.
Fifth, given the scarcity of numerous old descriptions, the lack of illustrations and/or measurements, the sterile condition of the material studied, or the presence of either male or immature female gonothecae, and the lack of knowledge of the cnidomes, only hardly reliable comparisons could be made. For example, there are several records assigned to H. delicatulum from South America and neighbouring seas, of which some are unrecognizable even in the presence of female gonothecae ( Blanco 1984a), while others, based on sterile material (e.g. Vervoort 1972), are simply unidentifiable.
Distribution in Chile. The former record is from Canal Vicuña (Galea 2007), while the present ones range roughly between 46° and 55°S.
World records. A subantarctic species possibly occurring from Marion Island to Patagonia.
10. The sample, preserved in ethanol, was received partly dried out. Upon examination in an aqueous solution, the gonothecal content appeared blurred. After a long storage in formalin, the air bubbles had completely disappeared, allowing its content to be observed properly during the present study.
Halecium Halecium delicatulum Halecium flexile Allman, Halecium Halecium Halecium Halecium Halecium balei Coughtrey, 1876 1888 humeriformis maximum modestum platythecum tristaniensis Fraser, sp. nov. sp. nov. sp. nov. sp. nov. sp. nov. 1911
……continued on the next page FIGURE 3. A: Halecium flexile Allman, 1888 —female (A1‒3) and male (A4) gonothecae, and comparison with the female of H. delicatulum Coughtrey, 1876 (B) and the males of H. balei Fraser, 1911 (C). D: Halecium humeriformis sp. nov. —female (D1‒3) and male (D4) gonothecae. E: Halecium maximum sp. nov. —female gonothecae (E), compared to the female of H. tristaniensis sp. nov. (G); male gonothecae (F). H: Halecium modestum sp. nov. — female (H1‒4) and male (H5) gonothecae. I, J: Halecium platythecum sp. nov. —female (I) and male (J) gonothecae. Scale bar: 500 µm.
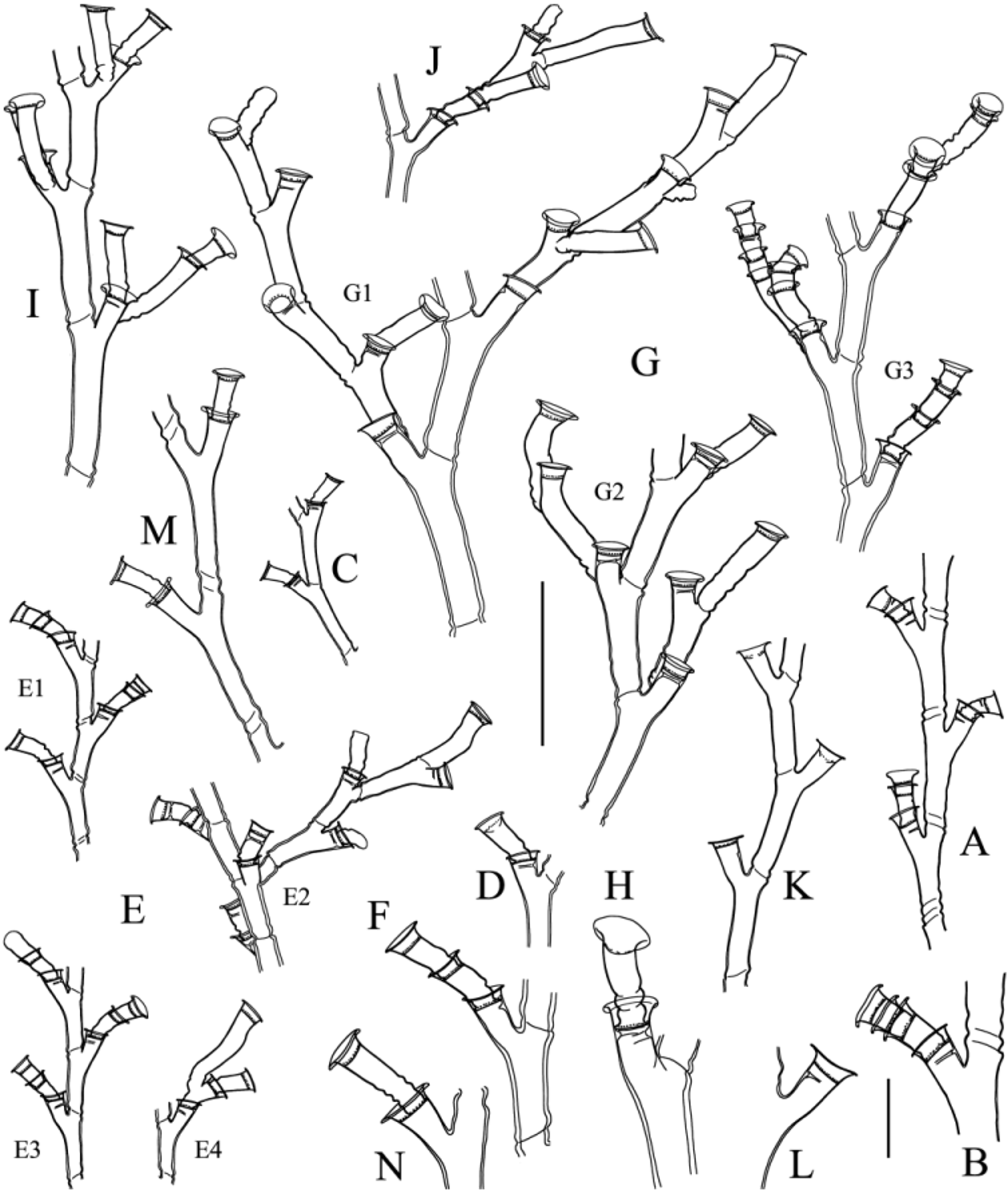



TABLE 4. Morphometrics of Halecium species discussed in the present study; dimensions in µm.
| Reference | Present study | Present study, Schuchert (2005) | Ralph (1958), as H. delicatulum | Present study | Millard (1977), as H. delicatulum | Present study | Present study | Present study | Present study | Galea (2010b), present study |
|---|---|---|---|---|---|---|---|---|---|---|
| Internode | ||||||||||
| - length | 415–575 | 360–690 | 500–870 | 620–1060 | 530–720 | 295–465 | 580–1110 | 465–900 | 735–930 | 455–560 |
| - diameter at node | 75–95 | 75–85 | 100–150 | 115–195 | – | 95–120 | 130–185 | 115–175 | 105–135 | 80–110 |
| Hydrophore | ||||||||||
| - length | 60–110 | 75–230 | – | 115–185 | – | 110–160 | 165–255 | 140–290 | 115–195 | 205–465 |
| Hydrotheca | ||||||||||
| - height | 30–40 | 50–65 | > 50–75 | 70–95 | 30–60 | 55–75 | 60–90 | 60–80 | 55–75 | 55–70 |
| - diameter at base | 95–105 | 90–115 | – | 115–140 | – | 100–120 | 125–155 | 130–155 | 130–140 | 110–125 |
| - diameter at rim | 125–145 | 165–190 | 140–190 | 200–230 | 170–200 | 145–185 | 200–240 | 215–245 | 205–240 | 160–200 |
| Hydranth | ||||||||||
| - tentacle number | Not seen | Not seen | – | ca. 22 | – | 19–22 | Not seen | 19–21 | Not seen | Not seen |
| ♂ Gonotheca | ||||||||||
| - length | 550–740 | Not seen | 700–1000 | 890–1010 | – | ca. 875 | 910–1205 | 865–975 | 730–1020 | Unknown |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Halecium flexile Allman, 1888
| Galea, Horia R., Schories, Dirk, Försterra, Günter & Häussermann, Verena 2014 |
Halecium flexile
| Millard 1977: 7 |
| Ralph 1958: 337 |
| Allman 1888: 11 |
Halecium delicatulum
| Coughtrey 1876: 299 |