
Cassidulus briareus, Souto & Martins, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4457.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:4A96289B-8C0A-4E5A-B2C0-B3D6E594B3CC |
|
DOI |
https://doi.org/10.5281/zenodo.5978151 |
|
persistent identifier |
https://treatment.plazi.org/id/0740DAFC-3232-4BA0-B18C-524BC68B5A11 |
|
taxon LSID |
lsid:zoobank.org:act:0740DAFC-3232-4BA0-B18C-524BC68B5A11 |
|
treatment provided by |
Plazi |
|
scientific name |
Cassidulus briareus |
| status |
sp. nov. |
Cassidulus briareus sp. nov.
( Figs. 1–2 View FIGURE 1 View FIGURE 2 , 10 View FIGURE 10 )
Type material. Holotype: Australia (Neuholland), collected by Salmin (probably Carl Ludwig Salmin), 18.36 mm TL (MP 1267 Holotype MNHWU). Paratype: same collection data as H, 18.20 mm TL (MP 1267 Paratype MNHWU).
Comparative material of other species examined. Cassidulus caribaearum? Lamarck, 1801 : possibly Caribbean Sea (MNHN-IE-2013-10590 [Syntype?]); Anegada, British Virgin Islands, TL 18–30 mm ( CASIZ
112633, 112638, 112683 A–B). Cassidulus infidus Mortensen, 1948b : Salvador–BA, Brazil, TL 7.5–13 mm (SMNH 4859 [H], UFBA 314, 757). Cassidulus mitis Krau, 1954 : Rio de Janeiro–RJ, Brazil, TL 26–46 mm (CASIZ 116110, MNRJ 3673–3674, ZUEC 11–12).
Diagnosis. Test arched in cross section, greatest height at apical disc. Anterior region of test very inflated. Paired petals with unequal columns of respiratory podia. Naked zone along oral midline developed and pitted. Phyllodes without occluded plates. Peristome and periproct transverse. Three interambulacral plates on each side of periproct. Basicoronal plate 5 narrow and elongated.
Etymology. Named after Briareus, the Greek God of violent sea storms, in reference to the turbulent taxonomic history related to the specimens described herein (see taxonomic history below).
Description. Test oval (TW 79% of TL), lateral edges straight, anterior and posterior margins round; anterior and posterior regions of aboral region inflated with greatest height at apical disc, transverse cross section domed; oral region concave along the midline of the anterior-posterior axis. Test measurements (TL * TW * TH in mm): 18.36 * 14.52 * 10.4 (H) ( Fig. 1A–B View FIGURE 1 ) and 18.2 * ca. 14.3 * ca. 10.4 (P) ( Fig. 1D–E View FIGURE 1 ).
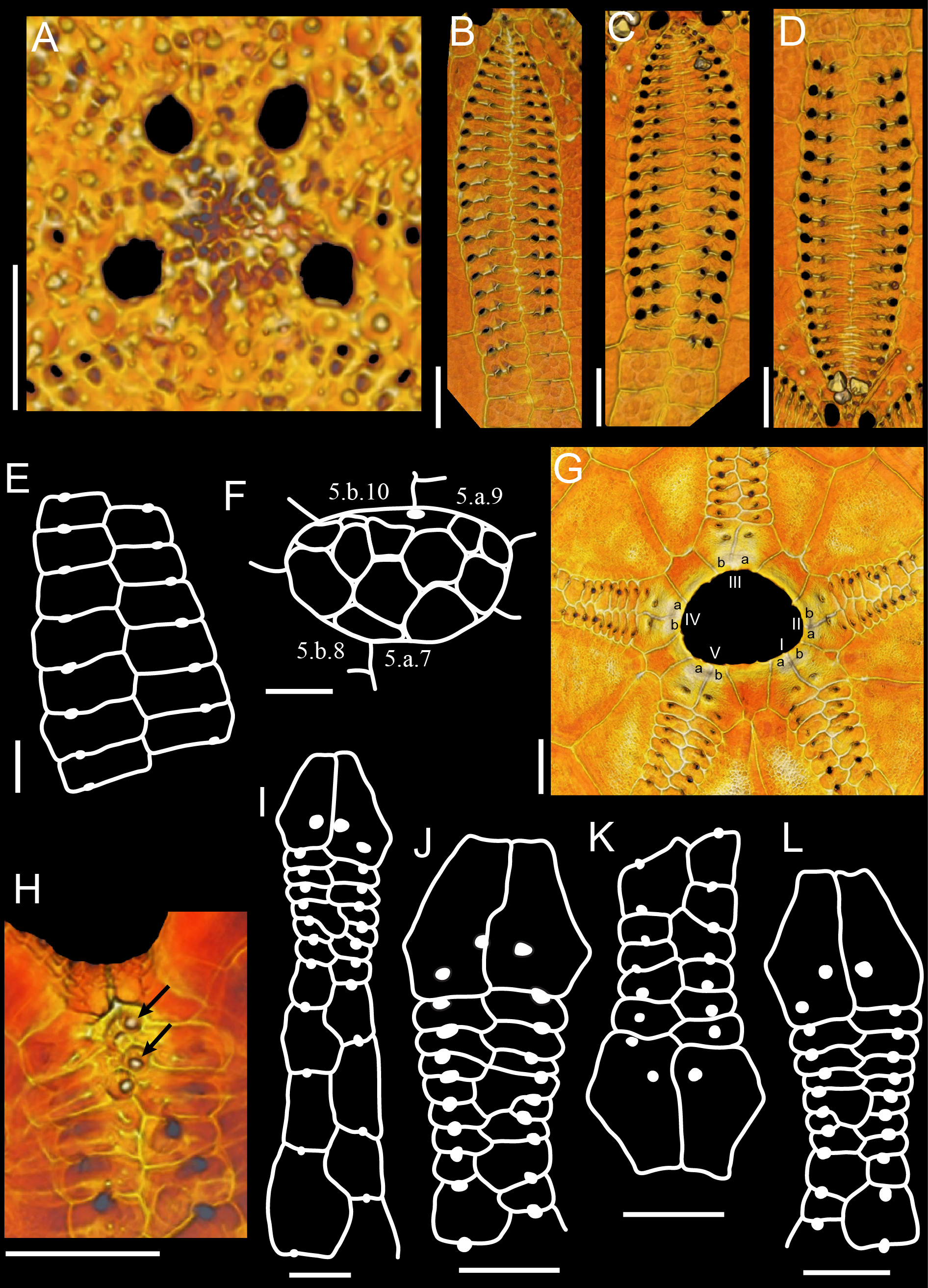
Apical disc anterior, monobasal, ca. 9.5% of TL, flat, with four gonopores on disc edge; hydropores abundant and spread across plate ( Fig. 2A View FIGURE 2 ). Anterior ocular plates between their adjacent gonopores; posterior ocular plates large and slightly posterior to gonopores 1 and 4; posterior region of apical disc slightly bulging towards interambulacrum.
Petalodium about 75% of TL. Petals roughly with same L and W, broad in the middle and narrow distally, but not closing ( Fig. 2B–D View FIGURE 2 ); inner and outer columns of respiratory podia bowed; poriferous zone narrow, pores slightly conjugated; outer pores elongated, inner pores round and smaller than outer. Columns a and b of posterior petals differ by 2 pore-pairs (number of respiratory podia in P: petal I, 21/23; petal V, 24/22), of anterior paired petals differ by 1–2 pore-pairs (number of respiratory podia in H: petal II, 21/20; petal IV, 18/20), anterior petal is equal (H: petal III, 21/21 respiratory podia). Primary tubercles present in poriferous zone; 3, sometimes 4 primary tubercles per petal plate. No occluded plates in petals. Ambulacra beyond petals increase 60–90% in relation to end-petal W; unipores in plates beyond petals: aboral plates wider than long, pores on suture, between adradial edge and middle of plate ( Fig. 2E View FIGURE 2 ), and oral plates about 2x longer than wide, pores on middle of plate suture ( Fig. 2I View FIGURE 2 ).
Phyllodes with unipores, with only one column of pores on each half (number of phyllopores per half: anterior phyllode 5–6, paired phyllodes 7–9) ( Fig. 2G–L View FIGURE 2 ). Plates sometimes unequal in size and shape; pyrinoid plate present on phyllode II, between plates 5 and 6 ( Fig. 2G, J View FIGURE 2 ). Columns slightly bowed proximally and W narrows down distally. Pores usually aligned in a uniform column. Buccal pores same size as phyllopores. Ambulacral basicoronal plates pierced by buccal pore and one per ambulacrum also pierced by a phyllopore in the sequence a, a, b, a, b from phyllode I to V ( Fig. 2G View FIGURE 2 ). Four to 5 sphaeridia in large and enclosed pits near buccal pores ( Fig. 2H View FIGURE 2 ).
Peristome anterior (34% TL from anterior region), deep (basicoronal plates extend further towards the inside of the test), slightly transverse (L 78–81% of W), pentagonal on the outside and subpentagonal on the inside ( Fig. 1C View FIGURE 1 ). Mouth opening in center of peristomial membrane. Bourrelets slightly developed as mounds mostly towards the inside of the peristome ( Fig. 10A–B View FIGURE 10 ). Cross-section of bourrelet with ca. 8 spines ( Fig. 10B View FIGURE 10 ).
Columns a and b of interambulacrum 5 with 7 and 8 plates between basicoronal plate and base of periproct, respectively, and 2 until adapical region of periproct (i.e., periproct is framed by 3 plates on each side). Interambulacral basicoronal plates 1 and 4 very narrow (much reduced and occluded in one specimen), 2 and 3 broadest, 5 intermediate in size ( Fig. 2G View FIGURE 2 ). Second and third oral plates on interambulacrum 5 are much longer than wide. Naked zone well-developed throughout midline of test on interambulacrum 5 and ambulacrum III (W 22– 25% of TW) ( Fig. 1B View FIGURE 1 ). Deep pits present in oral region, mostly on naked zone.
Periproct marginal and transverse (L 46% of W), beyond posterior petals; aboral plates form a prominent lip, oral and lateral plates do not bend inside the periproct ( Fig. 1E View FIGURE 1 ). Periproct framed adorally by plates 5.a.7 and 5.b.8, and adapically by plates 5.a.9 and 5.b.10. Periproctal membrane with 2 rows of 4 large plates; smaller plates scattered in aboral region near anus ( Fig. 2F View FIGURE 2 ). Anus opening aborally, on center of periproctal membrane.
Primary tubercles perforate and slightly crenulate. Oral primary tubercles with mamelon displaced in the opposite direction of the spine (usually anteriorly from center of bosses) and ca. 2.5x as large as aboral tubercles. Bourrelet spines curved and with thick tip, oral spines long and straight, aboral spines short and straight, spines on periproct thin, straight, and intermediate in size between oral and aboral spines. Miliary tubercles all over the test. Few tridentate pedicellariae on stalks (valves 220–225 mm L) on periproct. Because very little soft tissue was preserved on available specimens, ophicephalous pedicellariae and tube foot ossicles were not observed.
Remarks. Cassidulus briareus sp. nov. differs from its congeners (i.e., C. caribaearum , C. infidus and C. mitis ) ( Table 2) by having the anterior region of the test very inflated (vs. gradual height increase towards apical disc); a narrow and elongated basicoronal plate 5 (vs. squarish shape); a subpentagonal to triangular peristome, from the inside of the test (vs. subpentagonal to pentagonal); and a reduced number of plates framing the periproct (3 vs. 4 in the others [but rarely 3 plates on b column]). In addition, it differs from C. caribaearum by having an arched test in cross section of the adult (vs. triangular shape), shorter periproct, and more developed phyllodes with 2–3 more phyllopores than C. caribaearum ; from C. mitis by having a uniform row of pores in the phyllodes (vs. disorganized row in which some pores are displaced forming an apparent inner series), and from C. infidus by having the greatest height at the apical disc (vs. posterior to the apical disc).
Cassidulus briareus sp. nov. and its historical context within the genus Cassidulus . The specimens described herein were formerly identified as Cassidulus australis Blv. To our knowledge, Blainville did not describe any living species of Cassidulus and in his papers, he acknowledged Lamarck as author of C. australis (see Blainville, 1830, p. 192; 1834, p. 210). This species was described by Lamarck (1816) but later considered to be invalid ( Gray, 1855, p. 34; A. Agassiz, 1872 –1874, p. 153; Mortensen, 1948a, p. 209–210). Here we provide information on the taxonomic history of the genus Cassidulus that was pertinent in the decision to describe the new species.
Lamarck (1801, p. 348–349) described C. caribaearum together with the genus Cassidulus , without recording the repository of the holotype or the type locality of the species. The latter was assumed to be the Caribbean, given the name of the species and also because Lamarck mentioned Spanish Town (near Kingston, Jamaica) in a later publication ( Lamarck, 1816, p. 35). He (1801, p. 349) also made reference to Bruguière et al. (1827, pl. 143, fig. 8– 10), which includes illustrations of the specimen that, although of poor quality and lacking detail, captured the triangular shape of the test in cross section and the shape of the periproct and peristome.
In 1816 (p. 35), Lamarck described C. australis , based on specimens supposedly collected by Charles Alexandre Lesueur and François Péron during the Baudin Expedition (1801–1803) in Shark’s Bay, Western Australia (as “baie des Chiens marins, Nouvelle-Hollande ”). This description was slightly different from the description of C. caribaearum ; however, Lamarck mentioned that C. australis also occurred in the Caribbean and included in his synonymy list Bruguière et al. (1827, pl. 143, fig. 8–10), which means this species and C. caribaearum are synonyms. Again, he failed to record where the specimen was deposited.
Shortly afterwards, Lamouroux et al. (1824, p. 174) synonymized C. caribaearum with C. australis and redescribed the species. They also suggested re-naming the species C. richardi , in honor of the person who had collected the type of C. caribaearum , and mentioned that it was not clear that the specimen collected by Péron and Lesueur was the same as the one described by Lamarck (1816). Their description stated that the specimen was “rather convex above”, which matches the shape of Cassidulus briareus sp. nov., but the test shape in the illustration is more similar to that of C. caribaearum (triangular shape seen in large specimens).
All subsequent publications involving the genus Cassidulus (including the ones by Lamarck) accepted the synonymy proposed by Lamouroux et al. (1824), although any of the three names ( C. caribaearum , C. australis and C. richardi ) was used for the species (e.g., Eichwald, 1829; Blainville, 1830; 1834; Lamarck, 1837; 1840; Des Moulins, 1835 –1837; A. Agassiz, 1869; Mortensen, 1948b; Mooi, 1990b). The occurrence of the species in Australia was often disregarded even though Lamarck kept including this locality in his papers.
L. Agassiz & Desor (1847, p. 157) finally indicated that there was a specimen of C. australis (possibly the holotype) in the MNHN; and Mortensen (1948b, p. 209–210) suggested that the occurrence of the species in Australia was probably a result of “erroneous labeling”. His reasoning was that if the species occurred in Australia, more specimens would have been collected during the intervening 150 years; logic that makes sense for many taxa. This species, however, appears to be less common than expected.
Vadon et al. (1984) mentioned that they did not find the type of C. caribaearum in the MNHN and that Lamarck probably described the same specimen twice. The label retained the name used last, i.e., C. australis . However, Lamarck (1816) clearly mentioned that they were specimens from different localities. In addition, Lamarck’s descriptions of C. caribaearum and of C. australis were slightly different, although both were somewhat imprecise.
Possible type specimen of C. caribaearum at the MNHN. Following up on this debate, we believe that the vial identified as C. caribaearum in the MNHN ( Fig. 3 View FIGURE 3 , MNHN-IE-2013-10590 [Syntype?]; previously as EcEs 5040) might contain the type of one of the species described by Lamarck or perhaps even of both species. Two labels in the vial provide different identifications: one is identified as C. australis and the other (label apparently younger and written by A. Agassiz) is identified as Rhyncholampas caribaearum , with C. australis and Nucleolites richardi as synonyms. Also, both labels refer to the “Voyage de Péron & Lesueur 1801”, which indicates the material was from Australia rather than the Caribbean ( MNHN, 2017).
This vial contains broken pieces of two specimens as indicated by the fact that there are six complete phyllodes (one entire set and an additional broken phyllode). The following traits characterize them as Cassidulus : petal unequal in length (only one present, not sure if anterior or posterior; Fig. 3C, G View FIGURE 3 ), peristome transverse and subpentagonal (W 3.39 mm, L 2.18 mm), buccal pores present, sphaeridia in enclosed pits ( Fig. 3D View FIGURE 3 ), phyllodes with a single column of phyllopores per half ( Fig. 3D–F View FIGURE 3 ), naked zone developed and deeply pitted, mamelons of oral primary tubercles displaced from center. Additional characteristics are: posterior phyllodes and phyllode III with 5–6 pores, anterior paired phyllodes with 6–7 pores, plates beyond phyllodes longer than wide.
Basicoronal plate 5 seems more elongated than in specimens of C. caribaearum preserved in the CASIZ collection (CASIZ 112633, 112683 A) but not as narrow as in Cassidulus briareus sp. nov. ( Fig. 3A View FIGURE 3 ), and the number of phyllopores also overlaps both species. Therefore, identifying the broken pieces based on these traits is not reliable, nor would be any assertion that they belong to one or two species. Also, the illustrations in Bruguière et al. (1827, pl. 143, fig. 8–10) are very poor and only capture the overall shape of the specimen, making it impossible to determine if they correspond to the broken specimens in the vial at the MNHN.
Decision to describe a new species. According to the synonymy list provided by Lamarck (1816), C. australis is a subjective junior synonym of C. caribaearum . In addition, since C. australis was poorly described, it is impossible to determine if any subsequently collected Australian Cassidulus specimens are conspecific. The type specimen is probably lost, unidentifiable, or lacking diagnostic features, leading to much confusion in the literature regarding its validity. Therefore, it is not possible to determine that the material collected by Salmin is the same as Lamarck's C. australis , and we elected to describe a new species while at the same time stabilizing the nomenclature within the genus Cassidulus .
Prediction of the type locality of C. briareus sp. nov. C.L. Salmin was a trader in Hamburg, Germany, who collected and sold specimens during the 1860s–1870s without taking much care in recording details regarding sampling locality ( Holthuis, 2002). The only record we found in the literature documenting a collection of Australian echinoderms made by him is of holothuroids, collected in Cape York, Queensland ( Samyn et al., 2013). However, museum registries also indicate that he has collected marine fish in Victoria, suggesting that the specimens described herein could have been collected in other places along the Australian coast.
If “ C. australis ” was indeed collected by Péron and Lesueur in Australia, it is possible that C. briareus sp. nov. lived in the same region (i.e., Shark’s Bay, Western Australia).
Decision to designate a neotype for Cassidulus caribaearum Lamarck. As the type material of this species cannot be reliably identified and the illustrations provided by Bruguière et al. (1827, pl. 143, fig. 8–10) do not contain diagnostic characteristics, a neotype designation for C. caribaearum is necessary to objectively define this species name ( ICZN 1999, articles 75.1 and 75.3.4), and also to clarify the taxonomic status of the higher taxonomic ranks in which this species serves as the type species.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Cassidulus briareus
| Souto, Camilla & Martins, Luciana 2018 |
Cassidulus briareus
| Souto & Martins 2018 |
Cassidulus
| Souto & Martins 2018 |
Cassidulus
| Souto & Martins 2018 |
Cassidulus
| Souto & Martins 2018 |
Cassidulus
| Souto & Martins 2018 |
C. caribaearum
| ? Lamarck 1801 |