
Pseudosida szalayi Daday, 1898
|
publication ID |
https://doi.org/ 10.5281/zenodo.193382 |
|
DOI |
https://doi.org/10.5281/zenodo.5629379 |
|
persistent identifier |
https://treatment.plazi.org/id/03D987E7-FFCD-FFFE-0AF2-FC7262165C6E |
|
treatment provided by |
Plazi |
|
scientific name |
Pseudosida szalayi Daday, 1898 |
| status |
|
Pseudosida szalayi Daday, 1898 View in CoL
Daday 1898: 64−66, Fig. 33, a −d
Daday 1904: 111−112 ( Parasida )
Stingelin 1905: 335−337, Taf. 11, Fig. 1 View FIGURE 1 , 2 View FIGURE 2 ( Pseudosida szalayi ) Brehm 1953: 315−316, Abb. 83, 84 ( Pseudosida szalayi , P. bidentata ) Chiang & Du 1979: 99−100, Fig. 67 ( P. bidentata )
Idris 1983: 15, 18, Fig. 7 View FIGURE 7 , A −D ( P. bidentata )
Michael & Sharma 1988: 36−39, Text-figures 5, a −d, 6, a −e ( P. bidentata including var. szalayi ) Korovchinsky 1992: 67, 70, Figs 330−335
Korovchinsky 2004: 339, Fig. 134
Body measurements of specimens from the Lower Amur River basin are presented in Table 1 View TABLE 1 .
Parthenogenetic female. Body ovally-elongated with comparatively small head (27.6−35.1% of body length) often inclined down ( Fig. 1 View FIGURE 1 A, 2A). Rostrum large, directed more or less ventrally ( Fig. 2 View FIGURE 2 B). Eye large (6.9−10.5% of body length), situated near anterior or ventro-anterior head margin. Ocellus large, situated near base of antennules ( Fig. 2 View FIGURE 2 B). Antennules long (basal part 17.4−21.7%, the total length about 30% of body length) with aesthetascs on a small lateral prominence of their basal part, situated approximately in its middle or somewhat closer to base ( Figs. 1 View FIGURE 1 H, 2C). Basal part of antennules narrows distally, terminating in three apical prominences ( Fig. 7 View FIGURE 7 A). Long sensory seta, setulated distally ( Fig. 2 View FIGURE 2 C).
Swimming antennae long (51.9−67.1% of body length) with massive basipodite, having a triangular outgrowth on the posterior side and long, naked seta on dorsal side of its base ( Figs. 2 View FIGURE 2 A, 3G). Distal part of antennal basipodite bears rather long and stout spine on its outer middle side, small bifurcated prominence in front of upper branch, and large curved thorn together with large, wide horseshoe-shaped outgrowth dorsally ( Figs. 1 View FIGURE 1 B, C, 2D, 3F, I). Ventral corner of basipodital end may look rounded ( Figs. 1 View FIGURE 1 B, C) or with more or less developed outgrowth ( Figs. 2 View FIGURE 2 D, E, 3F). Inner side of basipodital distal end bears long feathered seta ( Figs. 2 View FIGURE 2 E, 3I). Proximal segment of upper two-segmented antennal branch (exopodite) with two small prominences proximally in front of bifurcated prominence of basipodite ( Figs. 1 View FIGURE 1 C, 2D). Distally this segment bears dorsally large spine, which may be of different size, straight or conspicuously curved, large sharpened outgrowth, and small prominence laterally ( Figs. 1 View FIGURE 1 D, E, 2F, 3A −E, I, 7B). Distal segment of this branch clearly articulates with previous one externally while internally these two segments seem more fused ( Fig. 2 View FIGURE 2 G). Distal segment bears small prominence proximally in front of large sharpened outgrowth of proximal segment (2G, 3D, 7B) while apically it has large spine, straight or curved, and small prominence near it ( Figs. 1 View FIGURE 1 F, 2H, I). All lateral setae of both segments armed uniformly with stout setules of swimming type while two longest apical setae armed proximally with similar setules but distally—with thin sparcely situated setules of sensory type (see Korovchinsky 1987). In lower three-segmented antennal branch (endopodite), second segment is most developed, long and stout, bearing apically one long seta (43.9−54.7% of body length in specimens from the Amurian populations) and strong spine. Distal segment of this branch is small and always bears three setae, one of which is almost as big as the one on second segment (37.7−47.0% of body length in specimens from the Amurian populations) (length ratio of these two setae 1: 0.9). In specimens from Lake Sur (Orissa, India) these setae are longer, reaching 59−71% of body length. Two other setae of this segment are smaller; two small spine-like prominences (rarely seen at the same time) are disposed near the setae bases ( Figs. 1 View FIGURE 1 G, 2J, 3H, 7C). Two large setae of lower branch occupy most ventral position; their proximal segments are naked, while distal segments armed with ventrally situated setules and terminally they have well developed hooks ( Figs. 3 View FIGURE 3 J, L, M, 7D). Two smaller setae of the branch armed distally with thin, sparsely situated setules of sensory type ( Figs. 3 View FIGURE 3 J, K) similar to those of apical setae of upper antennal branch. Formula of antennal setae: (4−7) − (9−11) / 0−1−3.
Mouth parts consist of maxillules (mx I), each with up to 11 densely situated feathered setae incurved forward, small maxillae (mx II) situated just near base of tl I ( Fig. 2 View FIGURE 2 L) and having short seta in the middle of their back side, and mandibles. Right mandible ( Fig. 8 View FIGURE 8 F) is comparatively narrow, with a group of three large thorns (pegs—Pe) posteriorly, diminishing in size ventrally, and four smaller ones near them. A row of five sharpened prominences (pales—Pa) goes along dorsal margin, while a row of 12 similar prominences (fishbones—Fb), three posteriormost of which are thinner and longer than others, is situated along ventral margin. Central part of molar surface with about ten diagonals (D) supplied with numerous pores. Left mandible ( Fig. 8 View FIGURE 8 E) is wider, with a group of pegs, consisting of two large and five small ones between and near them, at its dorso-posterior angle. From six to seven pales with finger-like prominences, then they seem to be fused all together and formed entire margin, go along the ventral margin of molar surface. These structures seem to form two or three rows, in inner of which only tips of pales are visible. A group of long fishbones near pegs, two or three of them are forked. Nine −ten diagonals, diminishing in size anteriorly, occupy most part of molar surface.
Shell with arched dorsal side, conspicuous dorso-posterior angle, and prominent, rather short (17.0−25.3% of body length) posterior margin smoothly connected with ventral margin ( Figs. 1 View FIGURE 1 A, L, 2A). Anterior valve margin is naked, lacks setae, which are situated lower on the ventro-anterior corner of valves, being directed outwards. These setae, four or five in number, are especially long and sit on developed conical bases ( Figs. 4 View FIGURE 4 C, D). The following six or seven marginal setae on the ventro-anterior margin appear directed forward and then setae become turned gradually inwards along inflexed ventral valve margin ( Figs. 4 View FIGURE 4 A, C). From seven to nine posteriormost setae on the ventro-posterior margin diminish in size dorsally, directing backwards and slightly outwards ( Figs. 1 View FIGURE 1 L, 4B). In total, there are about 45−50 feathered setae along valve margins. Posterior valve margin with an inner row of numerous submarginal spinules and clusters of spinules ( Figs. 1 View FIGURE 1 L, 7E, F).
Six pairs of thoracic limbs, all with epipodites. Their structure and armament are shown in Table 2 View TABLE 2 . Exopodite of tl I is comparatively narrow at its end ( Fig. 5 View FIGURE 5 A) while from tl II to tl VI it is widened terminally ( Figs. 5 View FIGURE 5 C, F). Endopodites from tl I to tl V are more or less clearly subdivided in four parts (segments), the proximal of which (I) is largest, bearing numerous filtering setae, while three terminal segments bear from four to seven such setae each. Terminal segment (IV) from tl I to tl V and subterminal segment (III) from tl II to tl V bear also outer setae each, similar with those of exopodite, of which the former one in especially long ( Figs. 5 View FIGURE 5 C, 6A, arrows). Large proximal segment of endopodite (I) of tl I also possesses terminally two naked setae, one of which is very long and another one is short (length ratio 6: 1) ( Fig. 5 View FIGURE 5 A). Endopodital filtering setae with very fine lateral setules, especially from tl II to tl V, and with a fascicle of more rough setules terminally ( Fig. 5 View FIGURE 5 D). A row of comparatively long rough setules along outer side of endopodite ( Fig. 6 View FIGURE 6 A). Gnathobase of tl I ( Fig. 5 View FIGURE 5 B) has outer row of 12−13 filtering finely setulated setae, ending with small naked seta (p) and inner group of the following structures: long two-segmented seta on distal corner with rough setules distally and fine setules proximally (I), comparatively large, curved downwards, distally setulated seta (i), and conical outgrowth between them (j). Gnathobases from tl II to tl V ( Fig. 5 View FIGURE 5 C) with row of 18−20 very finely setulated filtering setae, ending with one or two small naked setae (p) ( Figs. 5 View FIGURE 5 L, 6B), and distally with two setae of different type, one of which is long and two-segmented, resembling that one of tl I (I), and another one is strongly modified, being hooked terminally and denticulated laterally (J). Tl VI ( Fig. 5 View FIGURE 5 F) is small and strongly modified. Its exopodite has been reduced up to terminal plate, arming with six marginal setae and two large thorns near bases of two of these setae. Endopodite with seven similar setae and one thorn, situating laterally. Gnathobase with two long setae and three thorns of different shape.
L im b Exopodite (apical Endopodite Gnathobase Epipodite pairs + lateral setae)
I 7 + 4 (n 5–7 + 1) + (n 4) + (n 4) + (n 22 + 2 distal setae) n 12 + p + (I+j+i) * + II 7 + 4 (n 6 + 1) + (n 4 + 1) + (n 4) + (n 22) n 20 + p + (I + J) * + Postabdomen comparatively short (14.8−19.6% of body length) and high with long setae natatoriae (about 47% of body length) sitting on two separate bases ( Fig. 4 View FIGURE 4 G). Row of 10−12 clusters of anal teeth along each lateral side, mostly with 3−4, rarely 2, 5 or 6 teeth in each cluster; distalmost cluster is smallest ( Figs. 1 View FIGURE 1 I, J, 4E, 8C, D). Numerous groups and rows of spinules below the row of clusters of anal teeth (Figs, 8C, D). Groups of comparatively large spines near bases of postabdominal claws and anal opening ( Fig. 8 View FIGURE 8 A). Dorsally between bases of claws postabdomen bears terminal outgrowth ( Figs. 1 View FIGURE 1 I, J, 4E), which may be beak-shaped ( Figs. 1 View FIGURE 1 K, 4M, N, 5I, J), long ( Figs. 5 View FIGURE 5 G, H, K), or short (4E, H −L, 8D); setules on the outgrowth have not been observed. Postabdominal claws with three basal spines, two of which are long and one, proximalmost of them, is very small ( Fig. 4 View FIGURE 4 H −N, 8D). Two parallel rows of small denticles and spinules along outer lateral side of claws, a row of comparatively large denticles along their inner side, and numerous large spinules cover ventral side of the claws ( Figs. 8 View FIGURE 8 A, B). Body length 0.93−1.82 mm.
Gamogenetic female. All features as in parthenogenetic females, from which they differ in presence of dark, globular resting eggs in their brood pouches (diameter of eggs 0.22−0.24 mm). These eggs seem to possess rough shell and outside of brood pouch, they appear to be enclosed with transparent envelope. Body length 1.50−1.73 mm.
Male. Due to presence of only one adult male, proportions of most body structures have not been measured. Antennules long (46.0% of body length) with fused base and sensory seta ( Figs. 6 View FIGURE 6 F, G) and a row of densely situated thin, sharp spines along distal one third of the structure ( Figs. 6 View FIGURE 6 F, H). First pair of thoracic limbs (tl I) with large and complicated clasping organ ( Fig. 6 View FIGURE 6 C). It consists mainly of large hook and large bulbous structure in front of it, covered with numerous clusters of tiny spinules ( Figs. 6 View FIGURE 6 C, D). The latter one bears also long, stout thorn, while long, massive outgrowth attaches to lateral side of hook’s base. Filtering setae prove to be attached to back side of clasping organ ( Fig. 6 View FIGURE 6 E). Copulatory appendages tube-like and comparatively short ( Fig. 6 View FIGURE 6 I). Body length 0.80 mm.
Age, individual, and interpopulational variability. Juvenile females differ from adults in smaller body length (0.56−0.88 mm), fewer antennal setae (minimal number 3−7/0−1−3, more frequently 4−8/0−1−3), which sometimes are accompanied by setae’ rudiments, and fewer clusters of anal teeth (7−8 on each side of postabdomen). Juvenile males have body size 0.48−0.72 mm, shorter antennules (about 39.0% of body length) and short copulatory appendages.
In adult individuals, all measured structures show rather high individual variability (see Table 1 View TABLE 1 ), especially height of posterior valve margin and diameter of eye. The number of antennal setae and number of clusters of anal teeth seems not vary considerably while number of teeth in clusters is more variable. Two latter values often vary in one individual, being different on each side of postabdomen. The same is also true for number of filtering setae on terminal segment of tl I when, for example, there are six and seven setae on this segment in left and right limb, respectively.
The variability of dorsal postabdominal outgrowth is of special interest due to taxonomic importance of this feature. In specimens of one population, this outgrowth usually has uniform general shape, being small and short, beak-shaped, or long, but each of these types varies itself. This was clearly visible in specimens from the Lake Sur (Orissa, India) ( Fig. 1 View FIGURE 1 K) and especially clear in those from the Lower Amur River basin ( Figs. 4 View FIGURE 4 H −M). In one population from Bangladesh, there were specimens both with beak-shaped and long outgrowths and with comparatively short or long larger spines on the apical end of proximal segment of upper antennal branch.
Most of Amurian specimens were uniform, having small postabdominal outgrowth and not long and straight spines on the end of proximal segment of upper antennal branch but those from the pond near village Vinogradovka (site № 18) possessed a beak-shaped outgrowth and long, curved antennal spines ( Figs. 2 View FIGURE 2 I, 4N). Specimens from Yunnan ( China), India, and Sri Lanka had a beak-shaped outgrowth and not long, straight antennal spines, those from Lotus pond (Guandong, China), Nanjing ( China), and Kalimantan ( Indonesia) had long postabdominal outgrowth and long, curved antennal spines. On the other hand, these features in populations from the fish pond (Guandong, China), South Sumatra, and especially Bangladesh were mixed (long outgrowths and not long, straight antennal spines or in the latter case, long and comparatively short spines, beak-shaped and long outgrowths).
Remarks. This study is partly based on reinvestigation of original materials by E. von Daday from Sri Lanka ( Ceylon) (type specimens) and India. These materials were represented both by specimens on slides and in liquid samples. Part of specimens were in bad condition but others, including type specimens, have been preserved well enough to be analyzed. Most of their diagnostic features were accessible for the adequate comparison with those of specimens from other populations.
Having the original specimens, was possible to prove that Daday’s (1898) original description is incomplete and partly erroneous. Thus, in the antennule he described only two apical prominences in its base and naked sensory seta, in the swimming antennae-presence of sensory seta, not spine, on the outer side of basipodital distal end, in the postabdomen-presence of clusters with only three anal teeth and presence of only two basal spines on postabdominal claws. The ventral margin of valves was shown lacking setae.
Stingelin (1905) improved the description of the species considerably, describing correctly antennules, swimming antennae, postabdomen, postabdominal claws, and armament of valve margin. Posabdominal outgrowth of his specimens from Sumatra and Thailand ( Siam) was described as long, not beak-shaped as in Daday, and with tiny apical setules.
Being compared, the above mentioned descriptions by Daday and Stingelin look like concerned two different species. Only comparative investigation of sufficient material from different regions gives an opportunity to estimate the diagnostic features and their variability in species under consideration and conspecificity of representatives from different populations.
Differential diagnosis. The closest known species, P. bidentata , has not yet been studied in detail but the available materials ( Birge 1910; Korovchinsky 1992, 2004) allow concluding that it differs from P. szalayi in lacking of postabdominal terminal outgrowth, presence of an additional small seta on the terminal segment of lower antennal branch, and a prominence on the base of male’s antennules.
Type material. Two rather well preserved specimens studied by E. von Daday from type locality of Pseudosida szalayi (swamp near Lake Kalawewa, Sri Lanka ( Ceylon)) were reinvestigated by N.M. Korovchinsky in 1980 and selected as lectotype (adult parthenogenetic female with body length 1.26 mm on slide D XIII −44; II/P −48) and paralectotype (adult parthenogenetic female with body length 1.40 mm on slide D XIII −45; II/P −49). They are deposited in the Hungarian Natural History Museum (Budapest, Hungary) (see Forró & Frey 1982).
Another adult female from the same locality (slide D VIII −35; II/P −58) was undoubtedly collected and examined later and cannot to be considered type specimen.
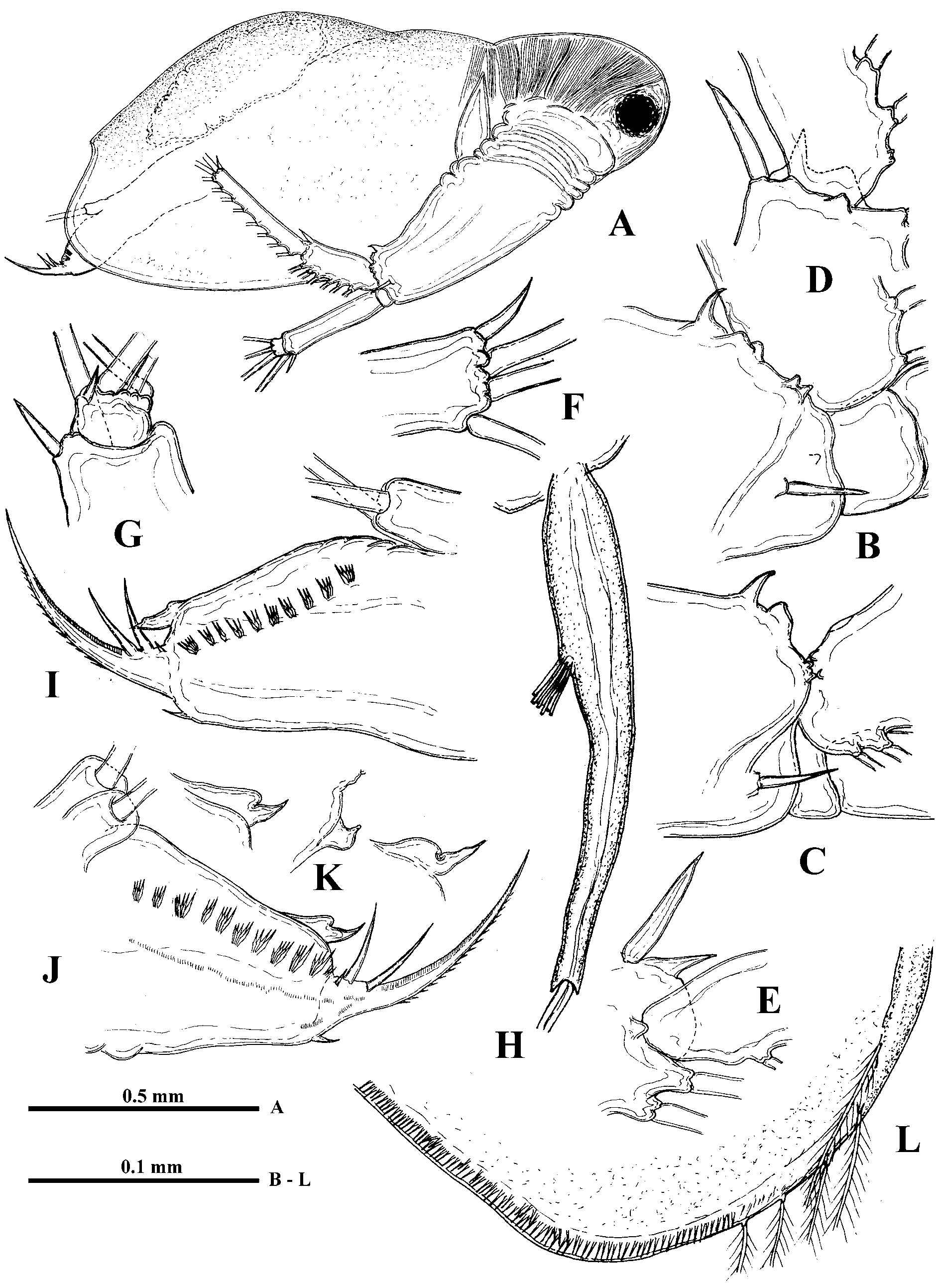
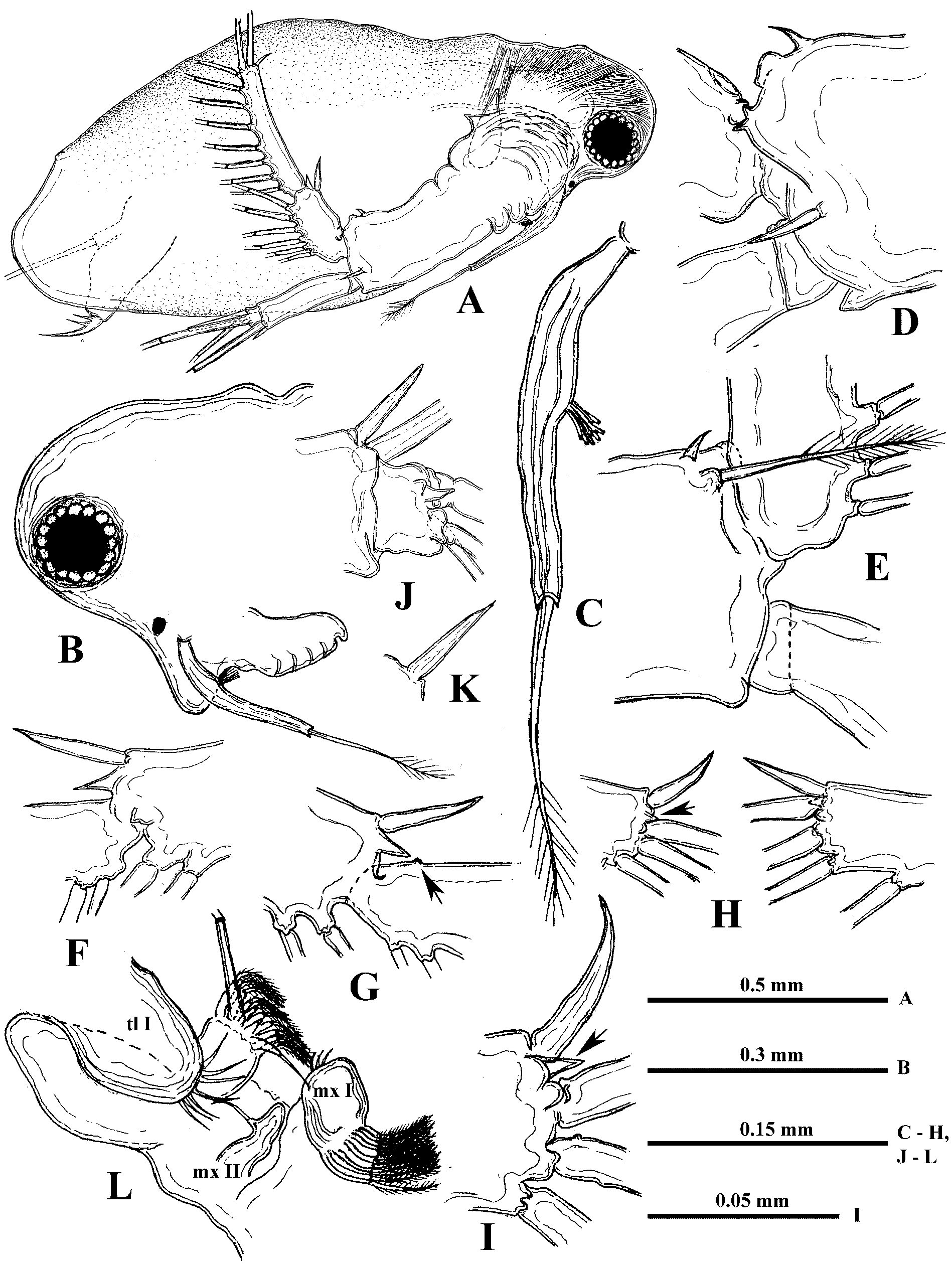
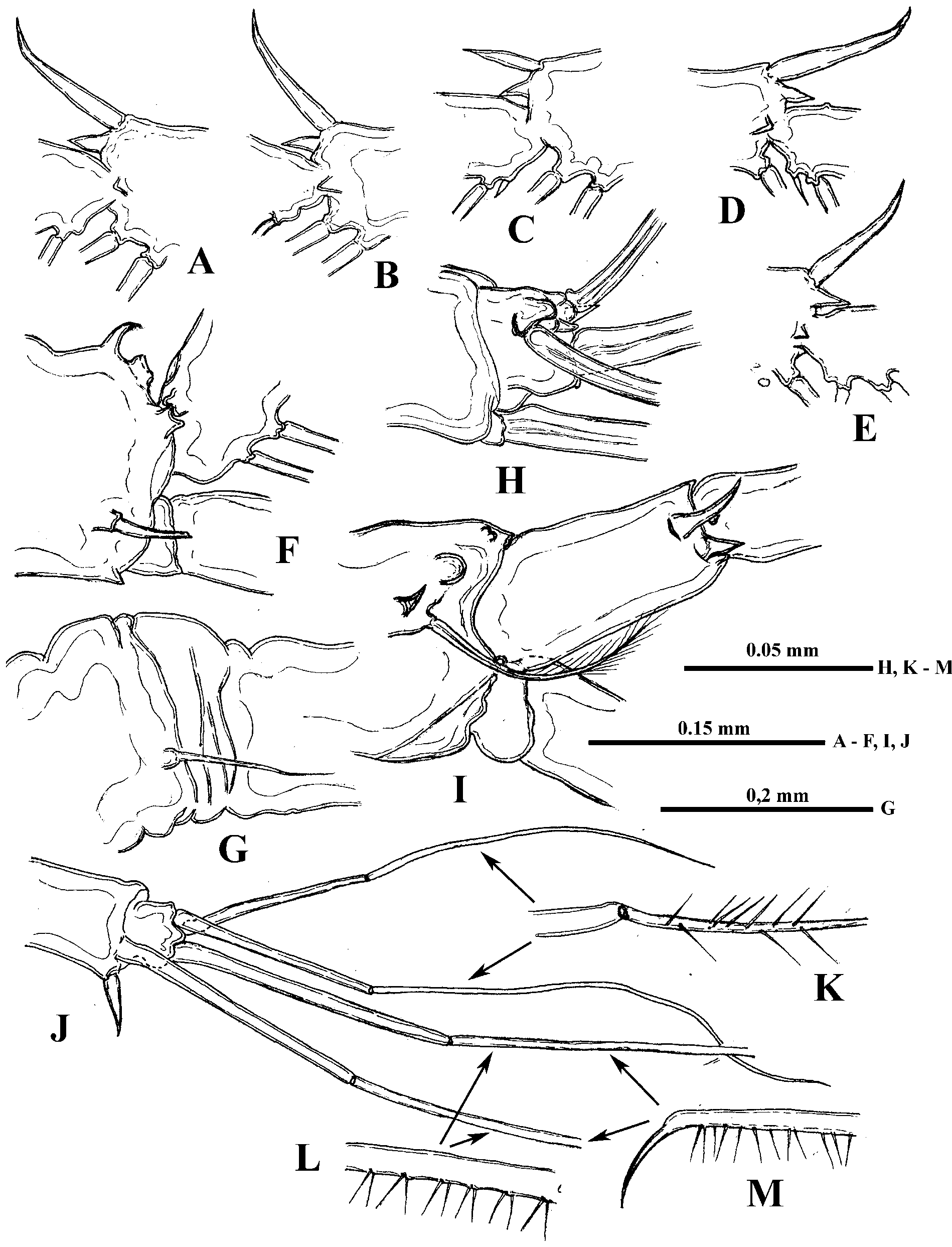
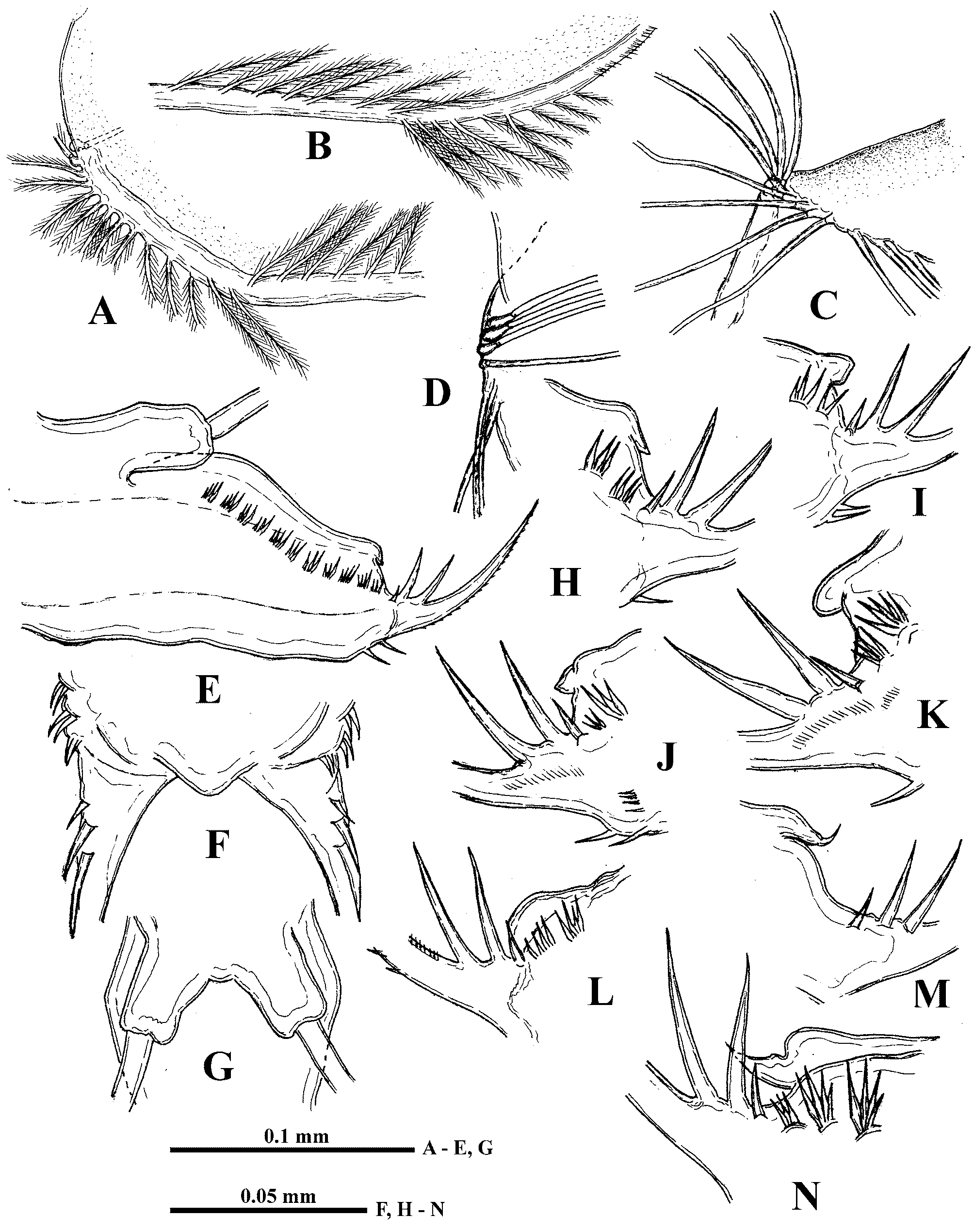
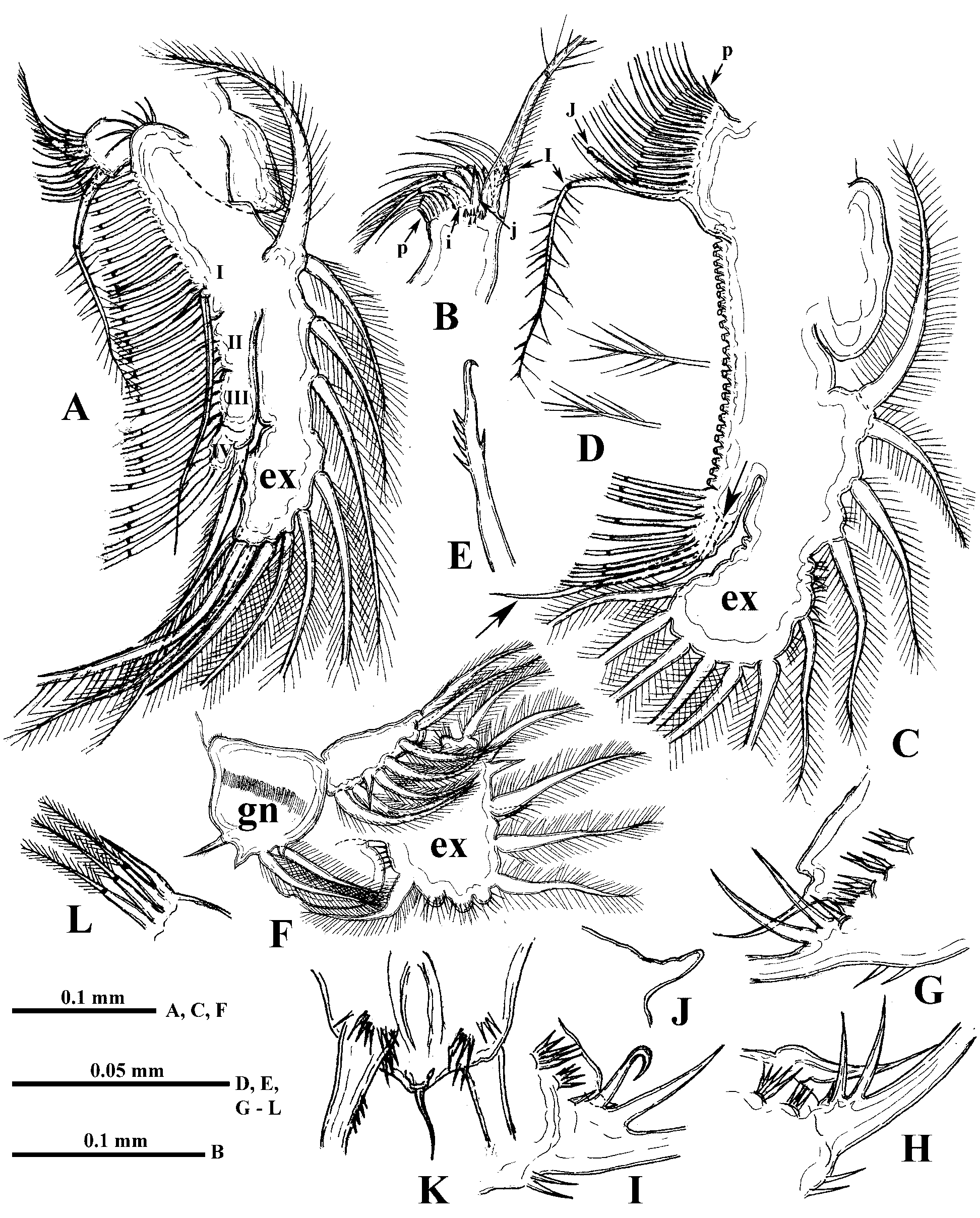
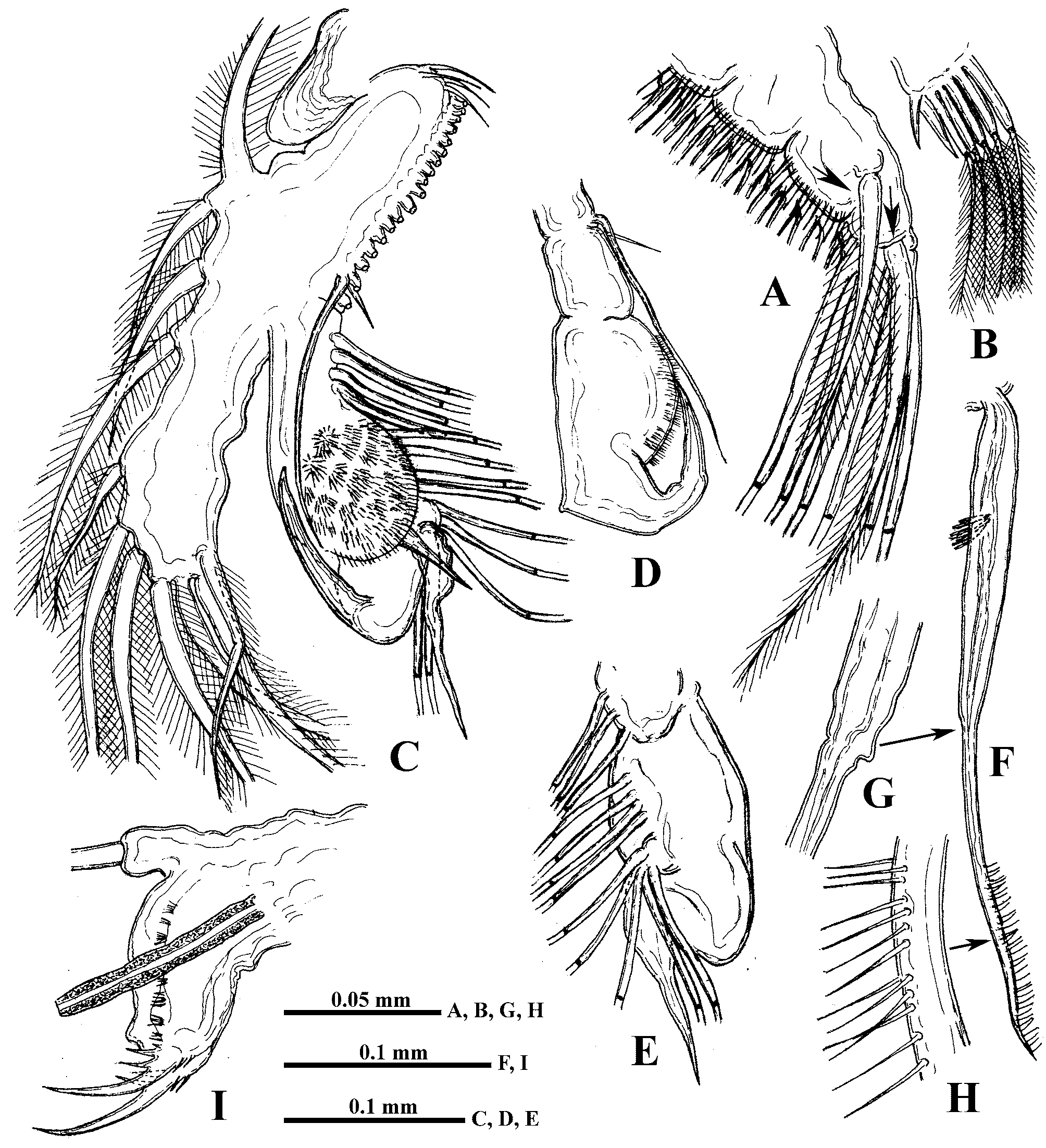
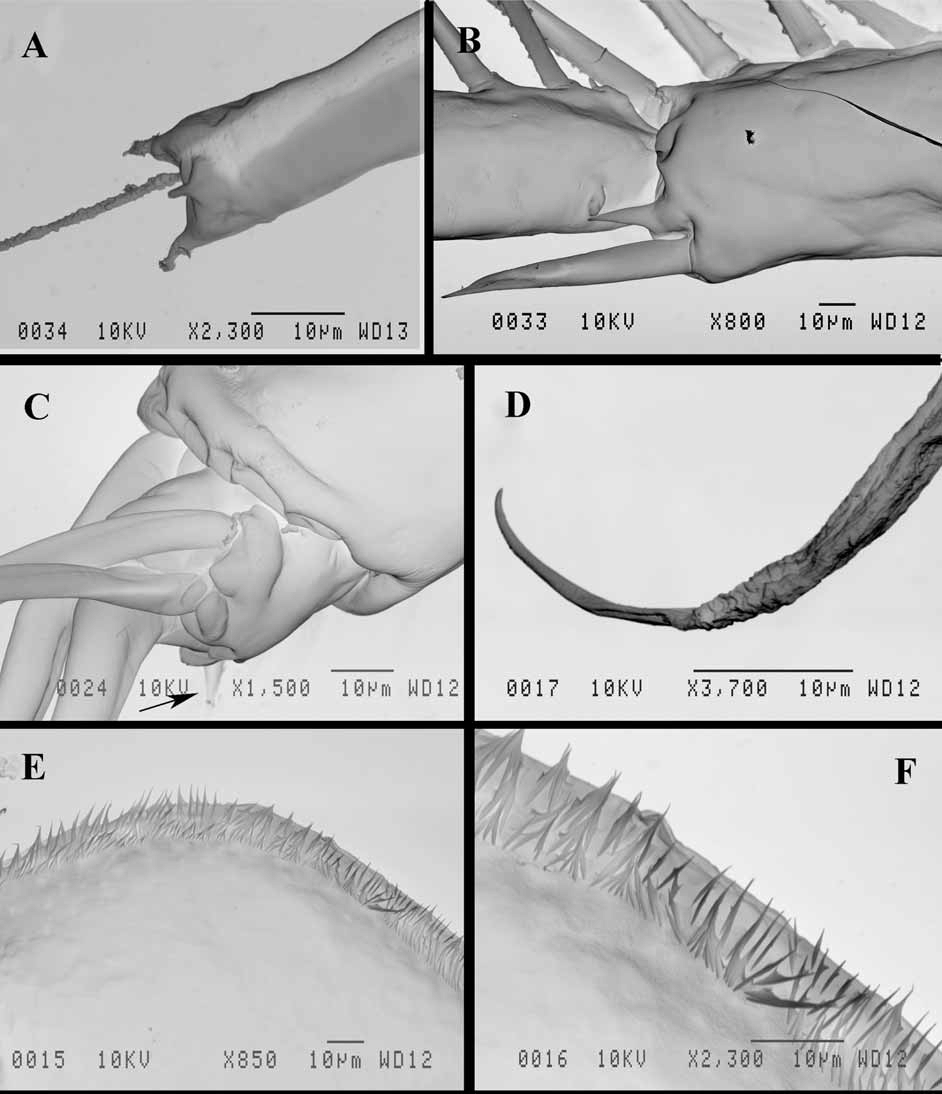
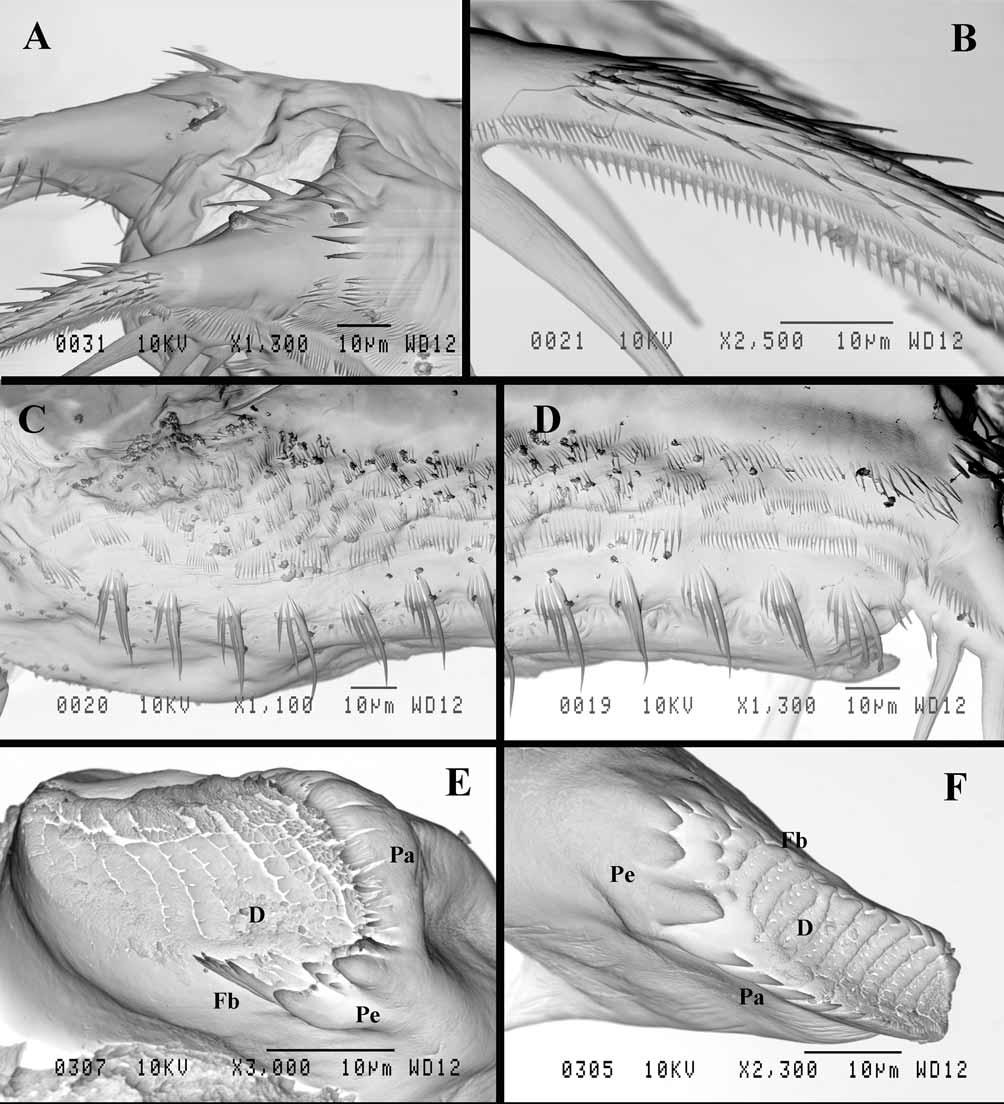
TABLE 1. Data on body measurements of the representatives of Pseudosida szalayi (n = 20) from the branch of Sinda channel (Lower Amur River basin) (in each column from top to down: range, mean, SD, CV).
| Body length | HL: BoL, % | DE: BoL, % | BAL: BoL, % | AnL: BoL, % | HPM: BoL, % | PoL: BoL, % |
|---|---|---|---|---|---|---|
| 1.20−1.82 | 27.6−35.1 | 7.4−10.5 | 17.4−21.7 | 51.9−67.1 | 17.0−25.3 | 14.8−19.6 |
| 1.38 | 31.2 | 9.4 | 19.4 | 59.8 | 21.6 | 16.1 |
| 0.17 | 2.23 | 0.72 | 1.30 | 4.10 | 2.20 | 1.03 |
| 12.3 | 7.2 | 7.7 | 6.8 | 6.9 | 10.3 | 6.4 |
TABLE 2. Data on structure and armament of thoracic limbs of Pseudosida szalayi from the branch of Sinda channel (Lower Amur River basin) (* see Abbreviations and Figure 5 for explanation).
| III 7 + 4 | (n 6 + 1) + (n 4 + 1) + (n 4) + (n 21) | n 20 + 2p + (I +J) + |
|---|---|---|
| IV 7 + 4 | (n 5 + 1) + (n 4 + 1) + (n 4) + (n 19) | n 20 + 2p + (I + J) + |
| V 6 + 3 | (n 4 + 1) + (n 3 + 1) + (n 3) + (n 15) | n 18 + p + (I + J) + |
| VI 5 + 1 | 7 | 2 + 3 outgrowths + |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.