
Cechetra bryki Ivshin & Krutov, 2018
|
publication ID |
https://doi.org/10.11646/zootaxa.4450.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:4EB291E9-75D9-46B3-8D77-B5D699D32AD3 |
|
DOI |
https://doi.org/10.5281/zenodo.5954788 |
|
persistent identifier |
https://treatment.plazi.org/id/03DA8796-FFBC-FFEA-BBEA-FCD4FDCBFC87 |
|
treatment provided by |
Plazi |
|
scientific name |
Cechetra bryki Ivshin & Krutov |
| status |
sp. nov. |
Cechetra bryki Ivshin & Krutov sp.n.
Holotype ♂, Nepal, Kathmandu [ Valley ], Mt. Pulchouki [ Phulchoki], 2770m [ca. 2500m], 16.vii.1990, ex. T. Haruta, E92-29, Cadiou Coll. BMNH (E) 2008-107 [ NHMUK] ( Fig. 18 View FIGURE 18 ).
Paratypes. 1 ♂, Nepal, Kathmandu, Godaveri, Phulchoki , 2400m, 26.v.1989, leg. [H.] Schnitzler, Cadiou Coll. BMNH (E) 2008-107 [ NHMUK] ; 6 ♂, India, Darjeeling, 3 miles Village, 29.vi.1986, leg. W. Thomas, 3, Cadiou Coll. BMNH (E) 2008-107 [ NHMUK] ; 1 ♂, Nord-Vietnam, Mt. Fan-si-pan, Cha Pa , 1700m, NN (22.15’N 103.46’E), 8.-29.v.1993, leg. Sinjaev & Simonov, [ex] Museum Witt, Sphingid genitalia slide No. 748, BMNH (E) 2002-25 [ NHMUK] ; 1 ♀ Burma, Mt. Victoria, Pakokku Chin Hills, 2600m, 2-31.v.1938, leg. G. Heinrich, Brit . Mus. 1938-689 [ NHMUK]; 2 ♂, 3 ♀, [ India,] Darjiling , leg. F. Möller, Rothschild Bequest B .M. 1939-1 [NHMUK]; 1 ♂, Bhutan, Buxa , Rothschild Bequest B.M. 1939-1 [ NHMUK] ; 2 ♂, 1 ♀, [ India,] Sikhim [ Sikkim], vii-ix, (from Fruhst. [ Fruhstorfer ]), Rothschild Bequest B.M. 1939-1 [ NHMUK] ; 1 ♂, [ India,] Sikkim, 26.iv.1888, leg. O. Møller, Collection H . J. Elwes, Rothschild Bequest B.M. 1939-1, Figured by Bernard D’Abrera [ d’Abrera ], 1986, in Sphingidae Mundi [ NHMUK] ; 1 ♂, China, Yunnan, Yuxi, Zhenyuan, Qianjiazhai , Ailaoshan Mt. , 2200 m, 1.viii.2011, leg. A. Kitching, 24°17'N, 101°15'E, ( BOLD sample id ARB00028595 About ARB ) [ CASIZ] GoogleMaps ; 1 ♂, Burma, Kambaiti , 2000 m, 1.vi.1934, [leg.] F. Bryk. Cechenena lineosa f. viridula Bryk 1944, type, coll. NHRS-TOBI 0 0 0 0 0 2260 [ SMNH] ; 1 ♂, Vietnam, Lao Cai Prov., Hoang Lien NP, Tram Ton , 1915 m, 22.3493723°N, 103.7704565°E, 8- 11.04.2010, leg. Papp L GoogleMaps ., Peregovits L. & Soltész Z., VN 2010PL_4 [ HNHM] ; 1 ♂, Nepal, S Annapurna Mt., S Banthanti , 2400 m, 28°21'N, 83°42'E, 25-26.vi.2008, leg. N. Ivshin ( BOLD sample id BC- Ivsh 01013), coll. N. Ivshin GoogleMaps ; 1 ♂, Nepal, S Annapurna Mt., Pothana , 2000 m, 28°19'N, 83°50'E, 5-6.vii.2008, leg. N. Ivshin ( BOLD sample id BC-Ivsh01014), coll. N. Ivshin GoogleMaps ; 1 ♂, Nepal, Nagarkot , 1990 m, 27°43'N, 85°31'E, 23.v.2012, leg. A. Bezborodkin ( BOLD sample id BC-Ivsh04735), coll. A. Bezborodkin GoogleMaps ; 2 ♂, N Vietnam, Lào Cai, Trạm Tôn Pass, N. side Fansipan Mt. , 1940 m, 16.v.–12.vi.2009, leg. V. Kroutov ( BOLD sample ids: BC- KVIMG4015-17 ) , coll. V.V. Krutov; 1 ♂, N. Vietnam, Lào Cai, Fansipan Mt. , 1940–2000 m, 25.iv.–15.v.2009, leg. V. Kroutov ( BOLD sample id BC-KVIMG4016-17), coll. V.V. Krutov ; 1 ♂, Burma, Kachin, Putao, Chudu Razi Hills , 25–28.vii.2008, leg. local collector ( BOLD sample id BC-KVIMG9968-17), coll. V.V. Krutov ; 7 ♂, N Vietnam, Lào Cai, Trạm Tôn Pass, N. side Fansipan Mt. , 1940 m, 16.v.–12.vi.2009, leg. V. Kroutov, coll . V.V. Krutov; 1 ♀, N Vietnam, Lào Cai, Trạm Tôn Pass, N. side Fansipan Mt. , 1940 m, 16.v.–12.vi.2009, leg. V. Kroutov, coll . V.V. Krutov; 2 ♂, China, Yunnan, Xinping, Mt. Ailaoshan , 2300m, vi.2017, leg. local collector, coll. V.V. Krutov.
Total: 39 paratypes.
FWL: holotype: 51mm
Morphologically, C. bryki is very similar to C. scotti and C. lineosa , but has a brighter green background with less prominent diagonal lines on the forewing upperside. The forewing ground colour is green (may be yellowishgrey, olive-grey or brownish-green in old or discoloured specimens) without the gradient in the green tone observed in C. lineosa . The diagonal lines of the forewing upperside are weakly differentiated from the ground colour, with some, particularly the most distal one, barely visible, unlike C. subangustata in which all eight dark green or brown diagonal lines are clearly visible. The outer edge of the black basal patch of the hindwing upperside extends along the veins. The female is similar to the male, but it is larger, with rounder forewings and with the black basal patch of the hindwing upperside with less pronounced sharp triangles extending along the veins.
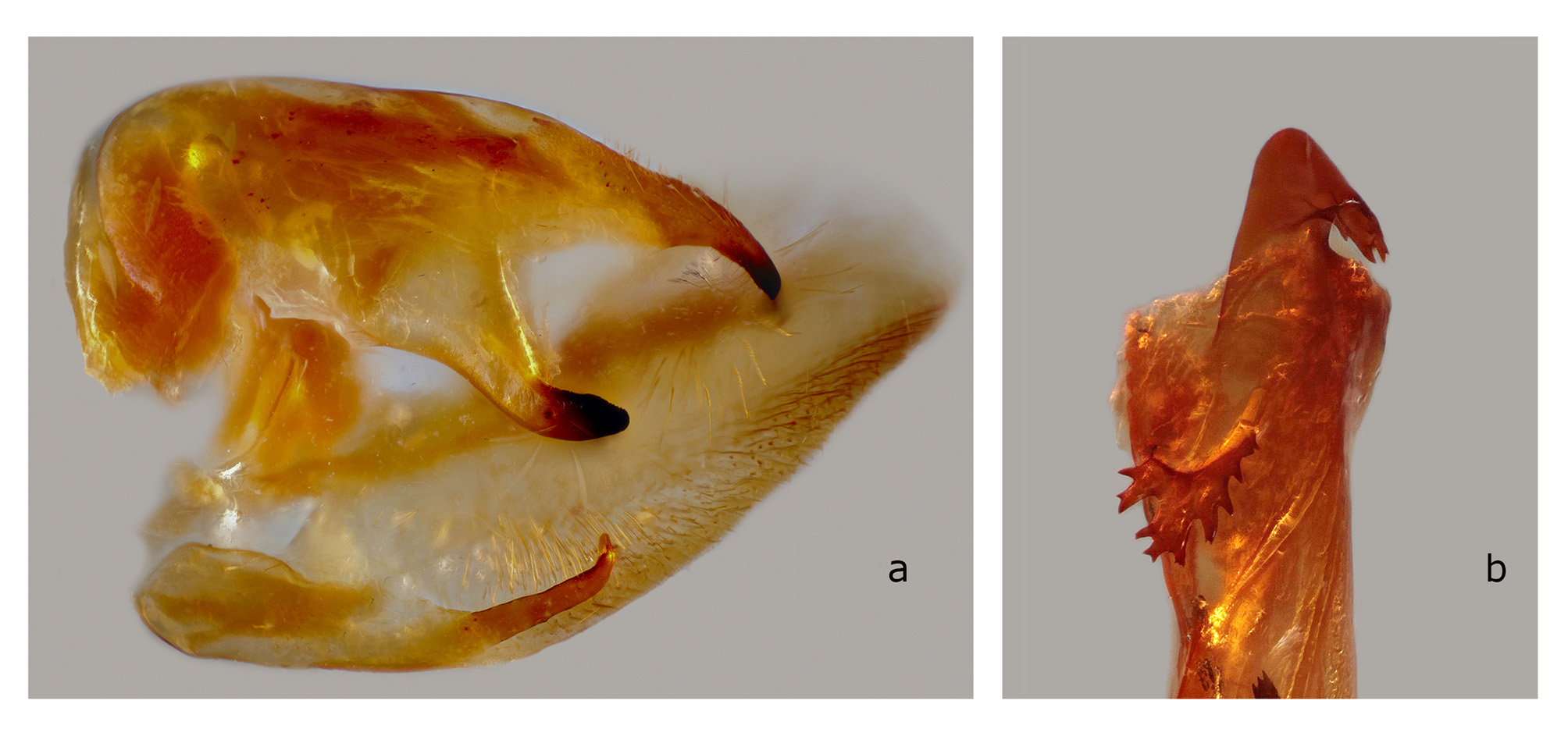
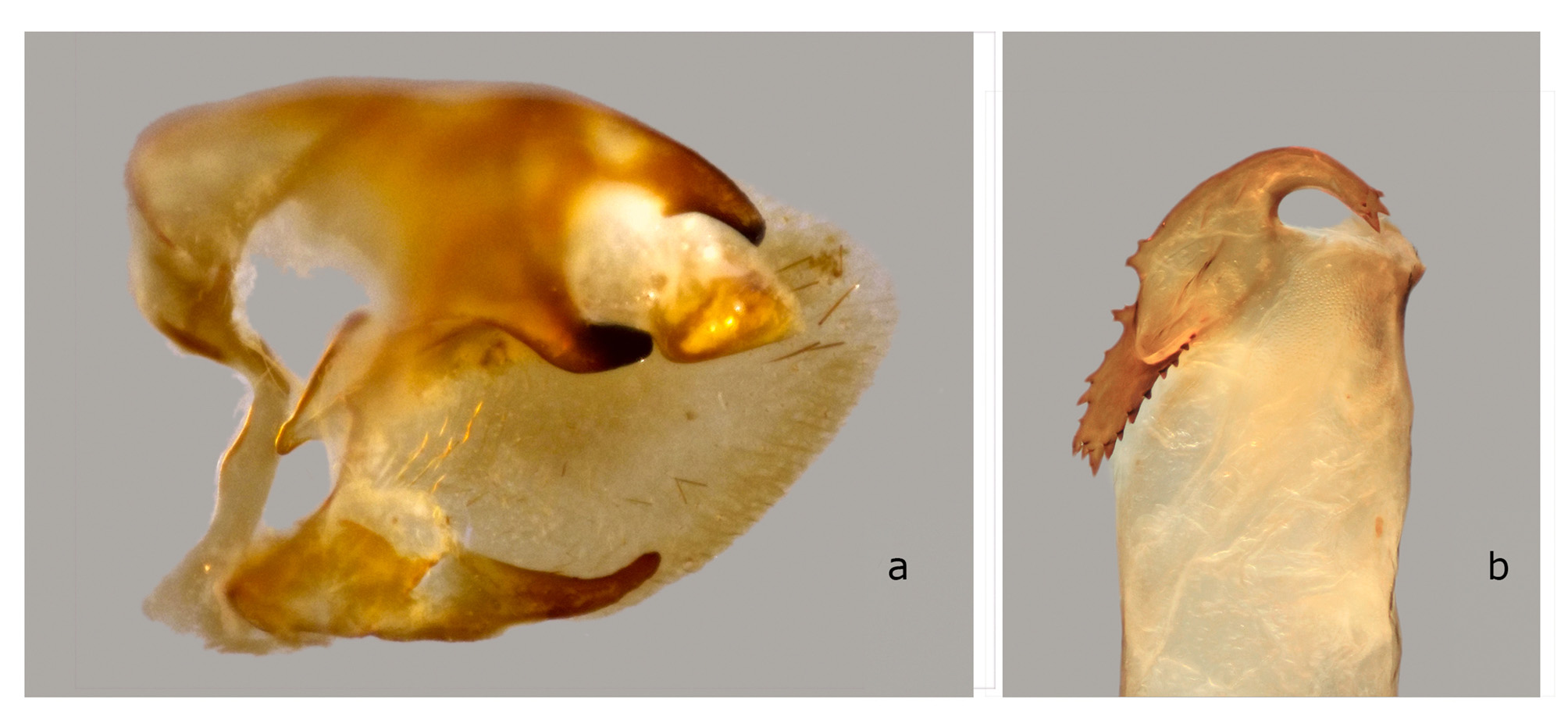
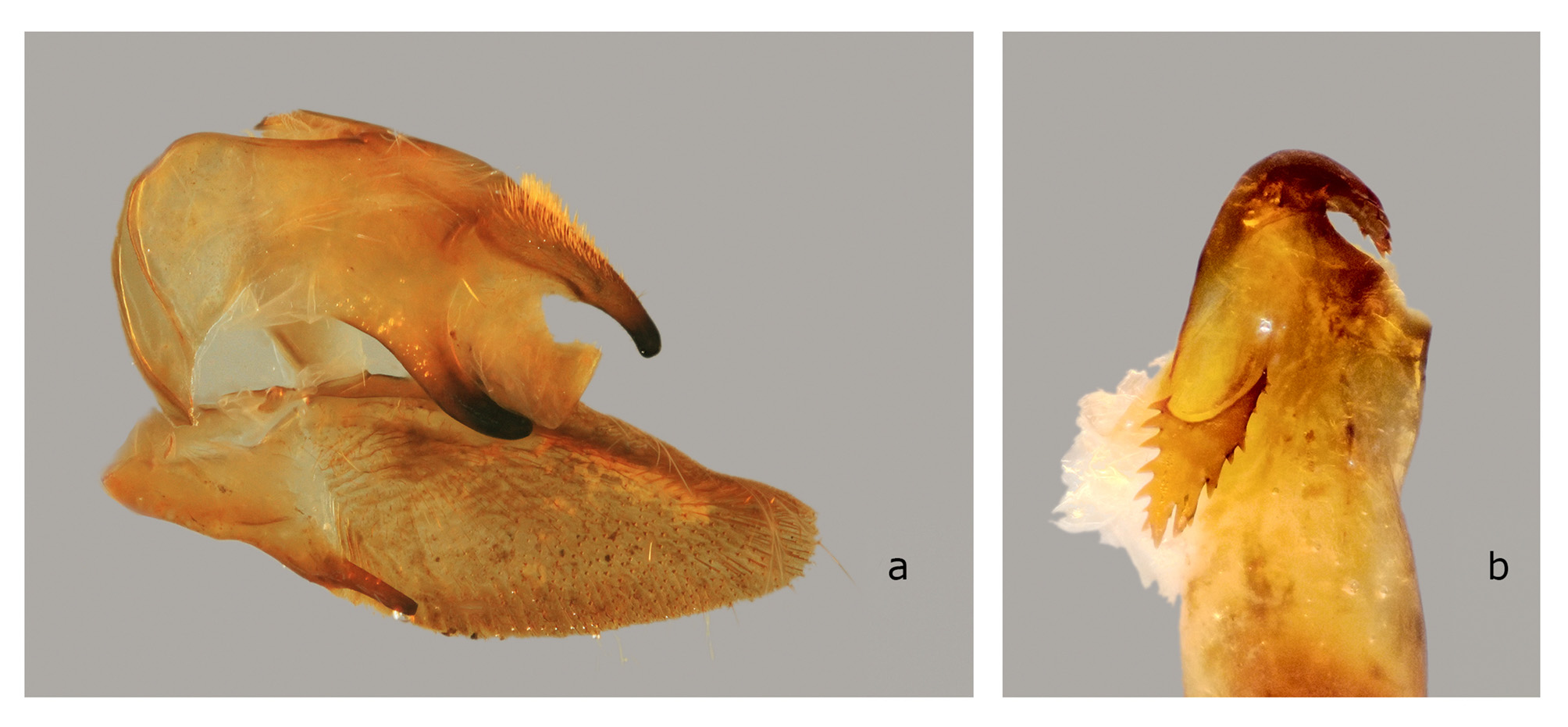
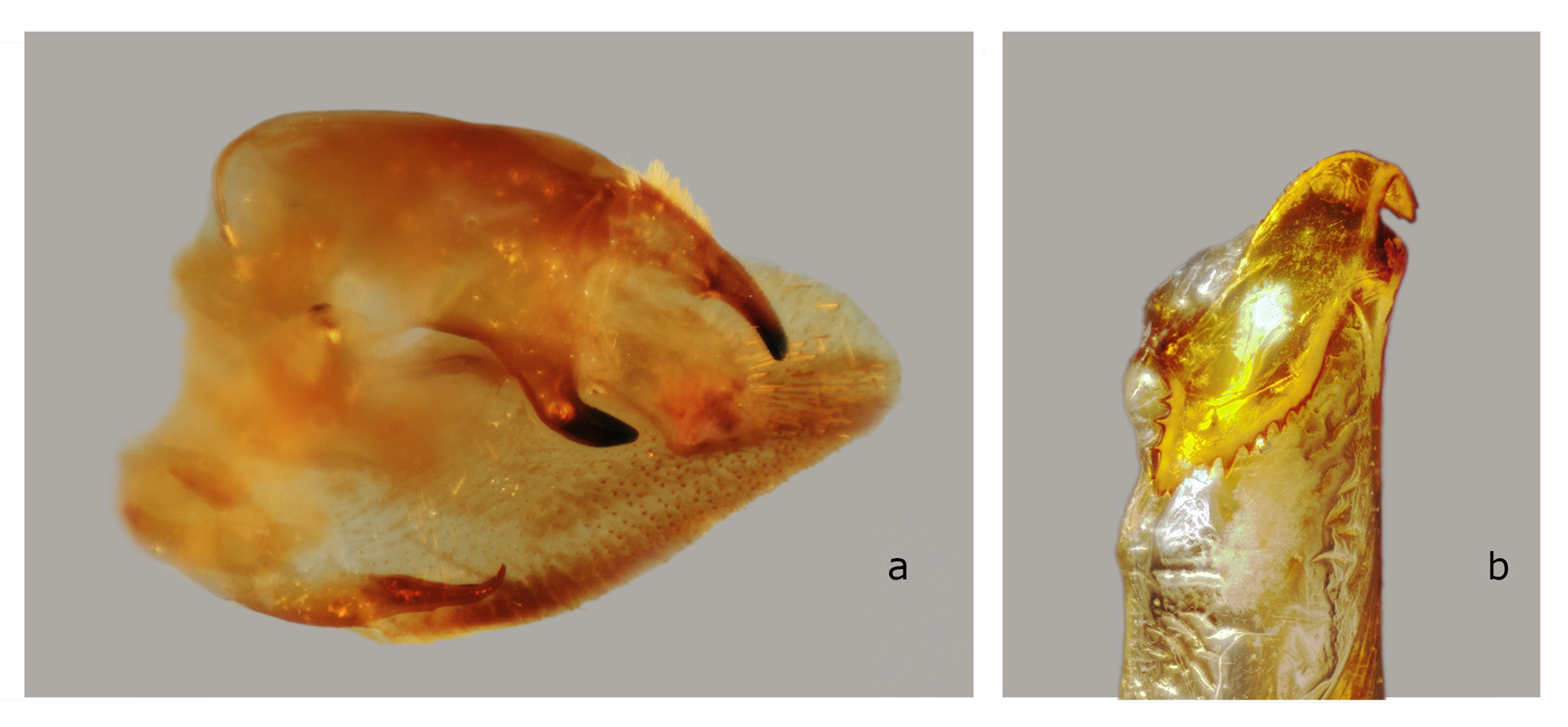
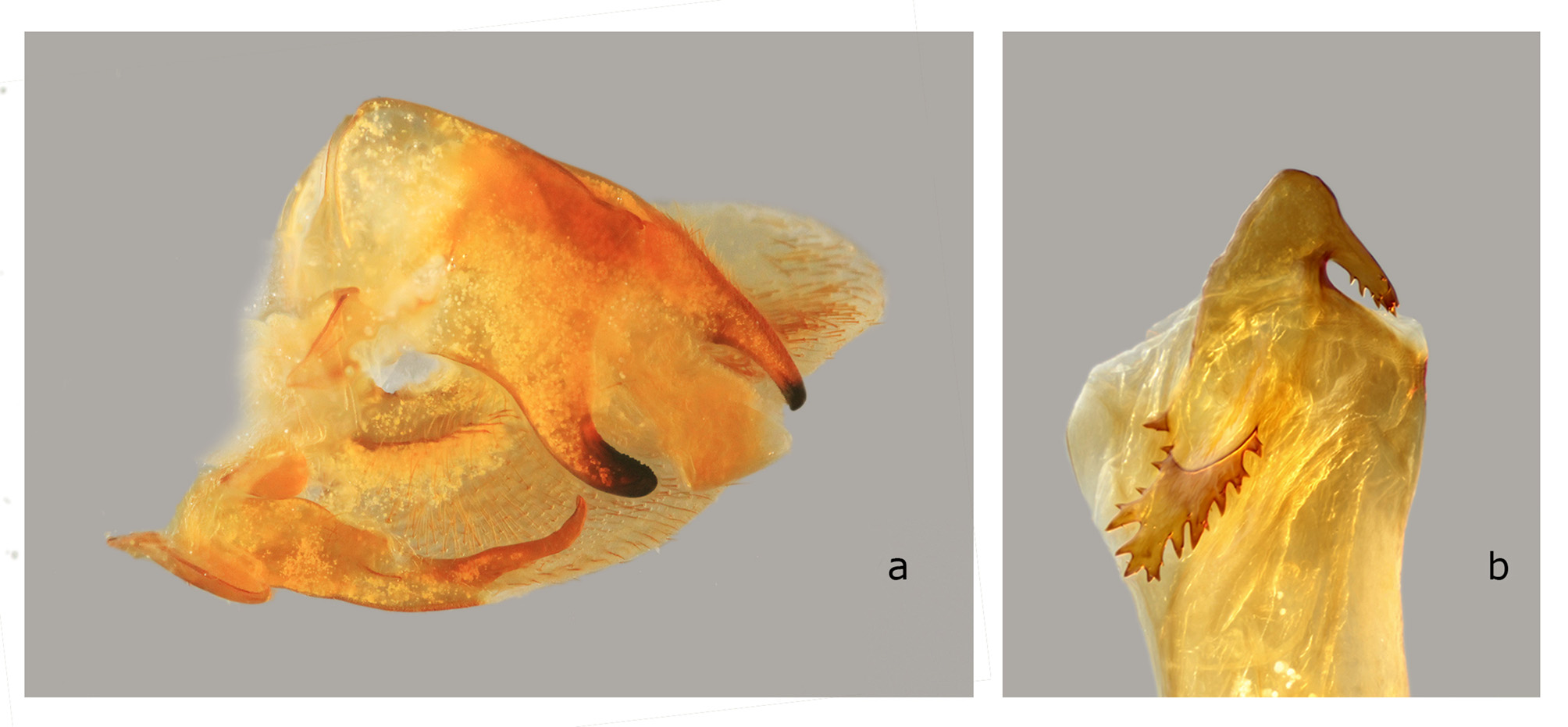
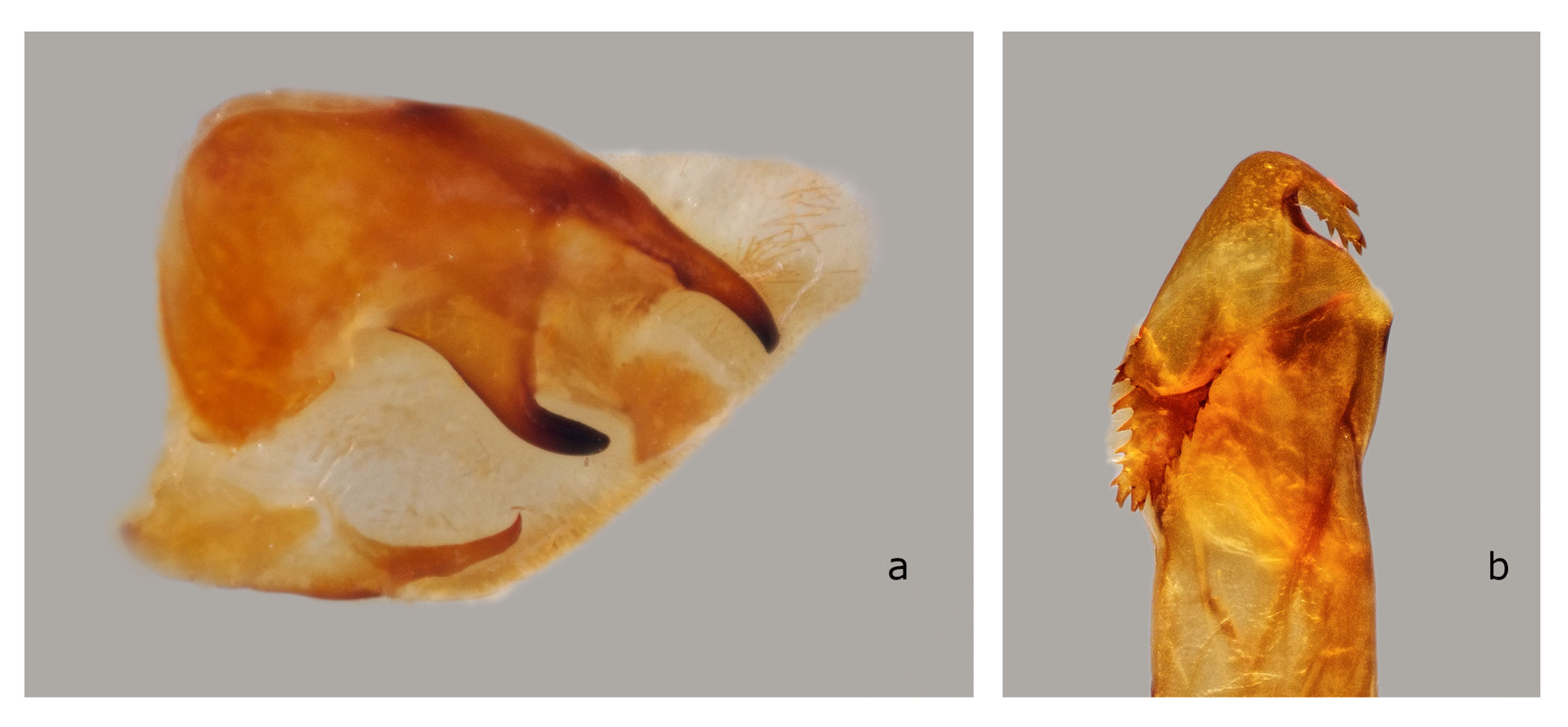
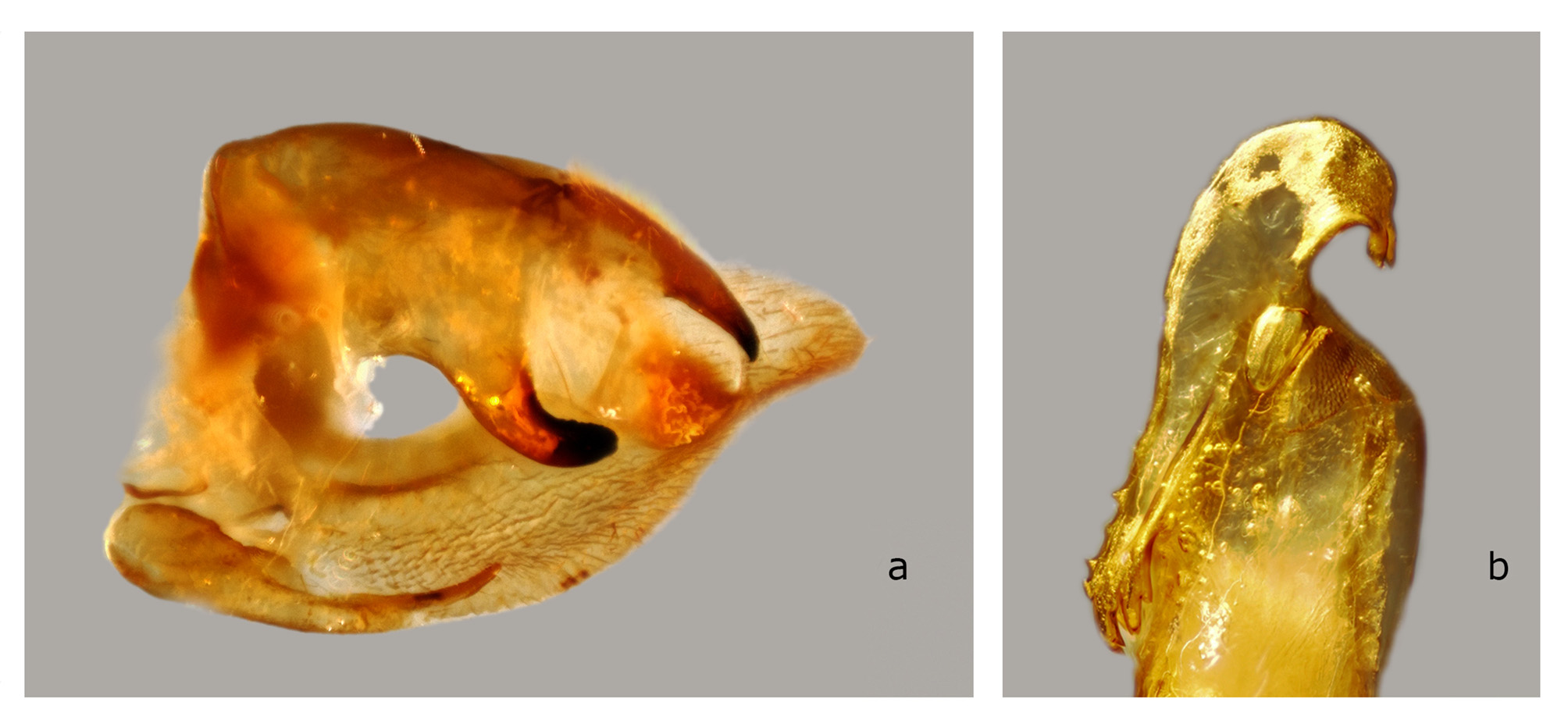
The structures of the male genitalia ( Fig. 21 View FIGURE 21 ) are within the range of variation of both C. lineosa and C. scotti ( Fig. 22–34 View FIGURE 22 View FIGURE 23 View FIGURE 24 View FIGURE 25 View FIGURE 26 View FIGURE 27 View FIGURE 28 View FIGURE 29 View FIGURE 30 View FIGURE 31 View FIGURE 32 View FIGURE 33 View FIGURE 34 ) and are very different from those of C. minor and C. pollux ( Fig. 35 View FIGURE 35 , 36 View FIGURE 36 ). The uncus lacks an acute sclerotized hook ventrally; the sclerotized process of the harpe is elongate, almost the same width over the entire length; and the margin of the basal plate of the phallus armature is deeply denticulate with a large apical hook distally (the basal portion of this hook is angled at about 30° to the phallus main axis). In male genitalia of the superficially similar but larger C. subangustata continentalis ssp. n. (described below) ( Fig. 37 View FIGURE 37 ), the uncus bears an acute tooth ventrally, the sclerotized process of the harpe is chunky, noticeably narrowing distally; and the phallus armature has a less denticulate plate and a short, small apical hook that is devoid of additional minute teeth on its surface (the basal portion of the hook is angled at about 45° to the phallus main axis). Generally, C. bryki has thinner and more elongate male genital structures than C. subangustata .
One mtDNA haplotype of C. bryki is shown in Fig. 40. It is common to all six examined specimens and the Chinese specimen found in the BOLD database. The translated polypeptide sequence differs from the other Cechetra species by one non-synonymic amino acid substitution.
Examination of the “type” of C. lineosa f. viridula convinced us that it is just a discoloured specimen of C. bryki . Another moth, in NHMUK, was illustrated by d'Abrera ([1987]: p. 205-206) as a “green form” of C. lineosa , but he did not give any locality data. We have examined this specimen, which proved to come from Sikkim, India, and found that it, too, is a specimen of C. bryki . We have thus included both these specimens in the paratype series of C. bryki .
Overall, the current known range of C. bryki encompasses areas in Northern India, Nepal, Myanmar, Southwestern China and Northern Vietnam at altitudes above 1900 m. It might also reasonably be expected to occur in the mountainous areas of Bhutan, Laos, Northern Thailand and Cambodia.
Currently, only one generation per year is known for C. bryki , from the end of May until early August.
The larval hostplants of C. bryki are currently unknown. The hostplants of the closely related C. lineosa were listed by Bell & Scott (1937) as Impatiens (Balsaminaceae) , Saurauia (Actinidiaceae) , Polygonum (Polygonaceae) and Vitis (Vitaceae) . All of these are members of early succession plant communities, which is typical for most hawkmoths of subfamily Macroglossinae . This explains, for example, why C. lineosa is a very common species in anthropogenic and disturbed plant communities. Such early succession communities at higher altitudes are usually microniches in late successional forests, known in the phytocenological literature as “gaps”, emerging due to rock collapse, windfalls and similar disturbances. Ecologically, C. bryki is likely to belong to the fauna of equatorial cloud forests and have a patchy distribution due to origin and lack of persistence of its habitats.
Etymology. The species is named after the Swedish entomologist, anthropologist and writer, Felix Bryk, who described C. lineosa f. viridula. We decided not to use the unavailable infrasubspecific name viridula but rather to propose a new name in accordance with the name availability criteria of the International Code of Zoological Nomenclature (ICZN 1999, 2012).











No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |