
Tubificoides mackiei, Kvist & Erséus, 2018
|
publication ID |
https://doi.org/10.11646/zootaxa.4433.3.10 |
|
publication LSID |
lsid:zoobank.org:pub:4E1F06CF-00B8-47E0-BD40-FA52B9C59F03 |
|
DOI |
https://doi.org/10.5281/zenodo.5998486 |
|
persistent identifier |
https://treatment.plazi.org/id/03DA87C5-9D57-FFD1-6482-6381FA60FD76 |
|
treatment provided by |
Plazi |
|
scientific name |
Tubificoides mackiei |
| status |
sp. nov. |
Tubificoides mackiei n. sp.
Figures 3–4 View FIGURE 3 View FIGURE 4
Tubificoides pseudogaster ; Brinkhurst & Baker 1979, partim?
Tubificoides pseudogaster ; Brinkhurst 1986, partim?
Tubificoides " pseudogaster I"; Kvist et al. 2010
Diagnosis. This species can be distinguished from congeners on the basis of the lack of hair chaetae and cuticular papillation, and the detailed morphology of the cuticular penis sheaths. In comparison with its closest known relative, T. pseudogaster , the new species is distinguished by its wider body at segment XI, its wider ental portions of the atria, its longer penis sheath, its lack of an outward-turned collar at the ental end of the penial tube, and its lower maximal number of preclitellar chaetae. In addition, the average COI distance between T. mackiei n. sp. and T. pseudogaster is 22.07% (see Kvist et al. 2010) and a total of 127 characteristic attributes ( i.e., molecular synapomorphies; see Sarkar et al. 2002) separate the COI sequences of these species ( Kvist et al. 2010).
Holotype. ZMBN 123768 (specimen ID CE21703), stained, whole-mount on microscope slide with coverslip, sexually mature specimen, collected by C. Erséus and M. Klinth, May 12, 2014; holotype COI sequence GenBank accession number MG652362 View Materials .
Type locality. Gunnarsholmen Island (inside of island, close to marina), Kragerö, Telemark, Norway ( 58°51'53"N, 09°24'42"E), 0.3–0.5 m depth, anaerobic, coarse sand and gravel. GoogleMaps
Paratypes. ZMBN 123769-123771 (specimen IDs CE21701, CE21702, CE21704); SMNH Type coll. 8955– 8957 (specimen IDs CE21705–CE21707), stained, whole-mounts on microscope slide with coverslip, all specimens fully mature (except CE21707), collected by C. Erséus and M. Klinth, May 12, 2014, from type locality. Paratype COI sequence GenBank accession numbers MG652360 View Materials , MG652361 View Materials and MG652363 View Materials .
Other material examined. SMNH 108985, SMNH 108986, and SMNH 108988 (specimen IDs CE199-3, CE2077, CE 3205 in Kvist et al. 2010), stained, whole-mounts on microscope slide with coverslip, one fully mature, one half-mature and one immature specimen, collected at Tjärnö Marine Biological Laboratory, Bohuslän, Sweden ( 58°52' 29"N, 11°08' 44"E) by C. Erséus, Sep. 2000, Oct. 2006, Sep. 2007; SMNH 108987 (specimen ID CE 3107 in Kvist et al. 2010), half mature specimen, collected at Hanhalsholme, Halland, Sweden ( 57°26' 58"N, 12°04' 02"E) by S. Kvist and M. Lindström, Sep. 2007; for genetic information, see Kvist et al. (2010).
Etymology. Named after the esteemed polychaete systematist, Andrew (Andy) S.Y. Mackie, for his contributions to annelid systematics, and for his hospitality, generosity and aid during the first author’s collection of numerous naidids (from various localities in Wales in 2007), including T. pseudogaster , which partly enabled the separation of the new species.
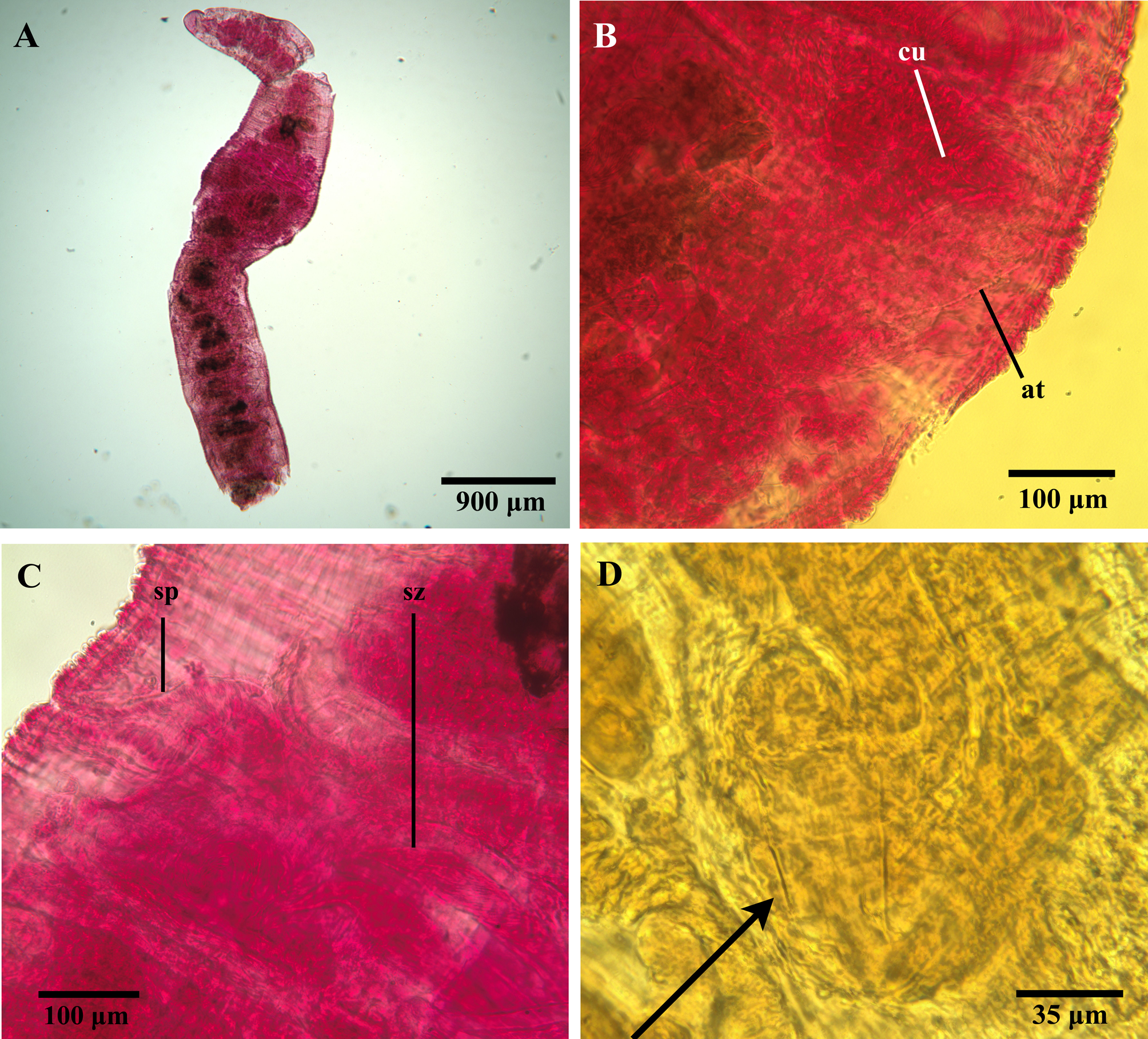
Description. All specimens incomplete, posterior ends used for DNA extraction. Holotype 4.52 mm long, with anterior 20 segments ( Fig. 3A View FIGURE 3 ), paratypes 1.7–4.1 mm long, with anterior 11–23 segments. Width at XI, 0.88 mm in holotype, 0.39–0.74 mm in paratypes; all specimens mounted and compressed. Holotype and all but one paratype sexually mature, other specimens in various stages of maturity. Prostomium rounded in most specimens, slightly more triangular in some, shorter than basally wide ( Fig. 3A View FIGURE 3 ). Cuticle smooth, cuticular papillae nowhere present. Clitellum present in X–XI in holotype and most paratypes, not developed in remaining specimens.
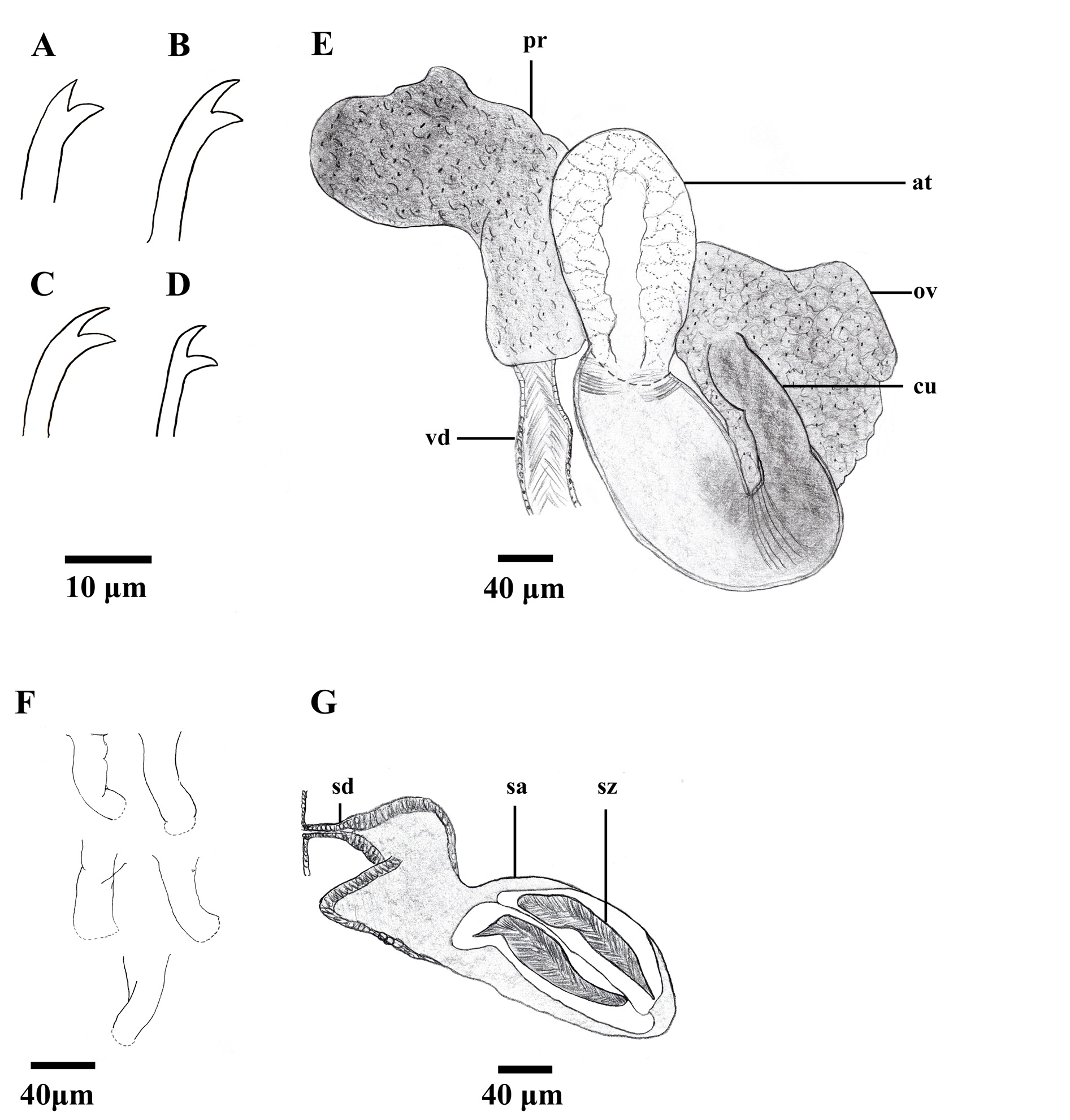
In preclitellar segments, dorsal chaetal bundles with (2)3–5 bifid crotchets ( Fig. 4A View FIGURE 4 ), 40–70 µm long. Bifids with teeth of about same length; upper tooth slightly thinner than lower. Postclitellar, dorsal bundles with 1–2(3) bifid crotchets ( Fig. 4B View FIGURE 4 ), 35–55 µm long; upper tooth longer and slightly thinner than lower. Preclitellar ventral bundles ( Fig. 4C View FIGURE 4 ) with (2)3–4 bifid crotchets (35–65 µm long); upper tooth thinner but about same length as lower. Postclitellar ventral bundles ( Fig. 4D View FIGURE 4 ) with 1–2 bifid crotchets (35–40 µm long), upper teeth slightly thinner than lower; teeth of about same length, and rather diverging. No ventral chaetae in segment XI in mature specimens. Dorsal bifid chaetae weakly sigmoidal, some ventral bundles with heavily bent, sigmoidal chaetae (sometimes broken); nodulus inconspicuous in most specimens, somewhat more developed in holotype. Spermathecal pores paired, located slightly above ventral chaetae in X. Male pores paired, in line with assumed ventral chaetae about halfway into segment XI.
Pharyngeal glands present in segments IV–V. Vas deferens ( Fig. 4E View FIGURE 4 ; vd) about 35 µm wide in holotype, full length not measurable in any specimen, but longer than 200 µm in one paratype; densely ciliated towards atrium insertion point. Prostate ( Fig. 4E View FIGURE 4 ; pr) rather large, but possibly compressed. Atrium ( Figs. 3B View FIGURE 3 , 4E View FIGURE 4 ; at) about 430 µm long, about 100 µm wide at widest point; tripartite with coarse cellular matrix in ental cap-like portion, finer cellular structure in mid-portion epithelium, and denser epithelium in muscular ectal portion. Atrium terminating in cuticular penis sheath with slight inward turn at ectal end, but still rather large terminal opening ( Figs. 3B View FIGURE 3 , 4E, 4F View FIGURE 4 ; cu), about 90 µm long, 30–35 µm wide at base. Soft tissue of penis protruding through terminal opening of penis sheath in holotype and most paratypes. Spermathecae ( Figs. 3C View FIGURE 3 , 4G View FIGURE 4 ; sp) extending from rather large but nonswollen pore, with narrower ectal duct ( Fig. 4G View FIGURE 4 ; sd) opening up into wider ampulla entally ( Fig. 4G View FIGURE 4 ; sa), spermathecae much compressed in all specimens and often pressed into surrounding segments; duct about 80 µm long, 40 µm wide at widest point, ampullae about 400 µm long, 150 µm wide at widest part. Spermatozeugmata rather stout and often coiled (170 µm long, 40 µm wide) ( Figs. 3C View FIGURE 3 , 4G View FIGURE 4 ; sz).
Distribution and habitat. Known from the southeast coast of Norway and west coast of Sweden, in fine sand and silt, as well as coarse sand and gravel; intertidal and shallow subtidal.
Remarks. Molecular data support that Tubificoides mackiei and the north European T. pseudogaster are closely related ( Kvist et al. 2010). The two taxa strongly resemble each other, both in general appearance and in some details of the chaetal morphology. In particular, Tubificoides mackiei and T. pseudogaster are similar in general body length from prostomium to segment XI, the lack of hair chaetae and cuticular papillation, and in the detailed morphology of dorsal chaetae ( Dahl 1960; Baker 1980; Takashima & Mawatari 1996). However, the two species can be distinguished on the basis of body width, e.g. using the width at segment X ( T. mackiei seems to be about twice as wide as T. pseudogaster in X [see Dahl 1960], although such a difference may depend on, or be exaggerated by, coverslip compression on a microscope slide), the shape of the atrium (wider ental portion in T. mackiei ), and the maximal number of preclitellar, dorsal chaetae (the reported [up to] six chaetae per bundle in T. pseudogaster [ Dahl 1960] are nowhere found in T. mackiei ). Also, the shape and size of the penis sheath (shorter in T. pseudogaster ), and the details of the penial tube (with an outward-turned collar at the ental portion in T. pseudogaster ) differs between the taxa. It should be noted that, with regard to the detailed morphology of the chaetae, in particular the length of the upper teeth in the dorsal bundles, Dahl’s (1960) original description and Baker’s (1980) re-description of T. pseudogaster report slightly different information. Dahl (1960) mentioned that the upper teeth in dorsal bundles are sometimes shorter than the lower teeth, whereas Baker (1980) suggested the opposite.
Dahl (1960) reported the following for T. pseudogaster : "In segment 9 the intestinal walls are thickened dorsally and laterally with a glandular structure". These structures are indicated in the scientific name for the species (Gr. pseudēs meaning false, and Gr. gastēr meaning stomach). All of our slide-mounted, non-juvenile specimens of both T. pseudogaster and T. mackiei display a feature very similar to that detailed by Dahl (1960), yet it is difficult to discern if the structures are of digestive epithelial origin or chloragogenic cells, and whether they are directly related to the intestinal tract or free in the coelomic cavity. Regardless, because both species seem to have this structure, it may be a synapomorphy for them (and perhaps additional, closely related taxa).
On the penis morphology of T. pseudogaster . Historically, delineations of species within Tubificoides have relied heavily on the shape and size of penis sheaths ( Brinkhurst & Baker 1979; Brinkhurst 1981, 1985, 1986; Baker 1983; Helgasson & Erséus 1987); this character also seems to separate T. mackiei from T. pseudogaster . Although the resolution of Dahl’s (1960) figure showing the penis sheath of T. pseudogaster is less than ideal, the language in her description is clear. Dahl (1960), when describing the penis sheath, states that: "Its length is about twice its breadth" and that "Distally the penial tube has an outward turned collar". In addition, the figures supporting Baker’s (1980) and Takashima & Mawatari’s (1996) re-descriptions of the species clearly show a "cone-shaped" penis sheath, with an outward turn at the ental end [referred to as "ental flange" in Takashima & Mawatari (1996)]. In order to aid future morphological identifications of T. pseudogaster (see also above regarding the DNA barcodes for this species) and owing to the low quality of the figures presented by Dahl (1960), we here present a photograph of the penis sheath from the lectotype of T. pseudogaster ( Fig. 3D View FIGURE 3 ). The structure is morphologically compatible with the detailed description by Dahl (1960), as well as both the text and figures presented in Baker’s (1980) re-description, and we hope that these new photographs, in combination with the genetic data, will mediate the difficulty of delimiting species that are morphologically similar to T. pseudogaster .
| SMNH |
Saskatchewan Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |