
SALMONIFORMES
|
publication ID |
https://doi.org/10.1111/zoj.12142 |
|
persistent identifier |
https://treatment.plazi.org/id/03DB6116-531E-A238-FB9A-D03621177A18 |
|
treatment provided by |
Marcus |
|
scientific name |
SALMONIFORMES |
| status |
|
SALMONIFORMES View in CoL View at ENA
ESOCOIDEI
Description
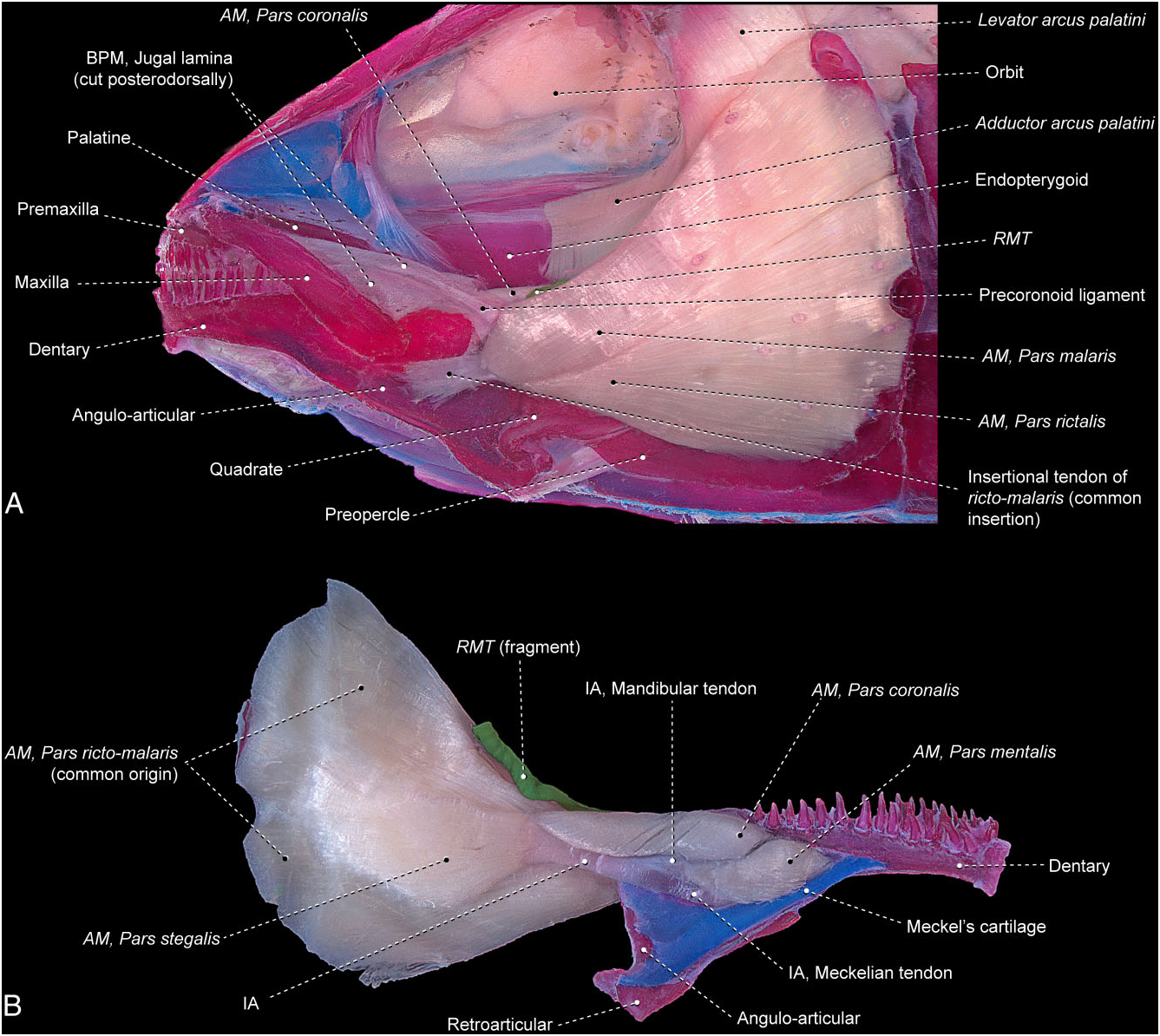
Umbra pygmaea ( Fig. 23 View Figure 23 )
The segmentum facialis arises from the preopercle, symplectic, and metapterygoid. This segment is mostly undivided along its extent, albeit with the three typical primary facial sections of the Teleostei still discernible, thereby indicating an incipient degree of differentiation amongst them. The stegalis is anteroventrally more obviously distinguishable because of its shorter muscle fibres that arise from the metapterygoid and insert on the intersegmental aponeurosis. This aponeurosis is divided anteriorly into a meckelian tendon that inserts on the coronomeckelian, and a mandibular tendon, which serves as the site of origin of the segmentum mandibularis. The lateral-most fibres of the segmentum facialis are superficially arranged into two bundles anteriorly. The dorsal bundle presumably corresponds to the malaris and the ventral bundle to the rictalis. These two external bundles converge anteriorly to a tendon that continues forward to its insertion on the lateral surfaces of the dentary and angulo-articular. The medial-most fibres of the rictomalaris retain an attachment on the intersegmental aponeurosis in conjunction with the fibres of the stegalis. The retrojugal lamina has a well-differentiated ligament running from the coronoid process of the lower jaw to the anteromedial region of the maxilla, but no muscle fibres of the adductor mandibulae are associated with this ligament. This ligament apparently does not correspond to any of the ligaments most commonly running within the buccopalatal membrane across the Teleostei (see Datovo & Vari, 2013) and is herein termed the precoronoid ligament.
The segmentum mandibularis is completely divided into a coronalis and a mentalis. Posteriorly, the coronalis extends beyond the limits of the lower jaw to arise from the dorsal margin of the intersegmental aponeurosis and from a mandibular raphe that is shared with the inner fibres of the ricto-malaris. Anteriorly, the coronalis inserts on the posterodorsal portion of the dentary. The mentalis originates from the mandibular and meckelian tendons and inserts onto the angulo-articular and dentary.
The ramus mandibularis trigeminus nerve passes between the ricto-malaris laterally and the posterior portion of the coronalis medially.
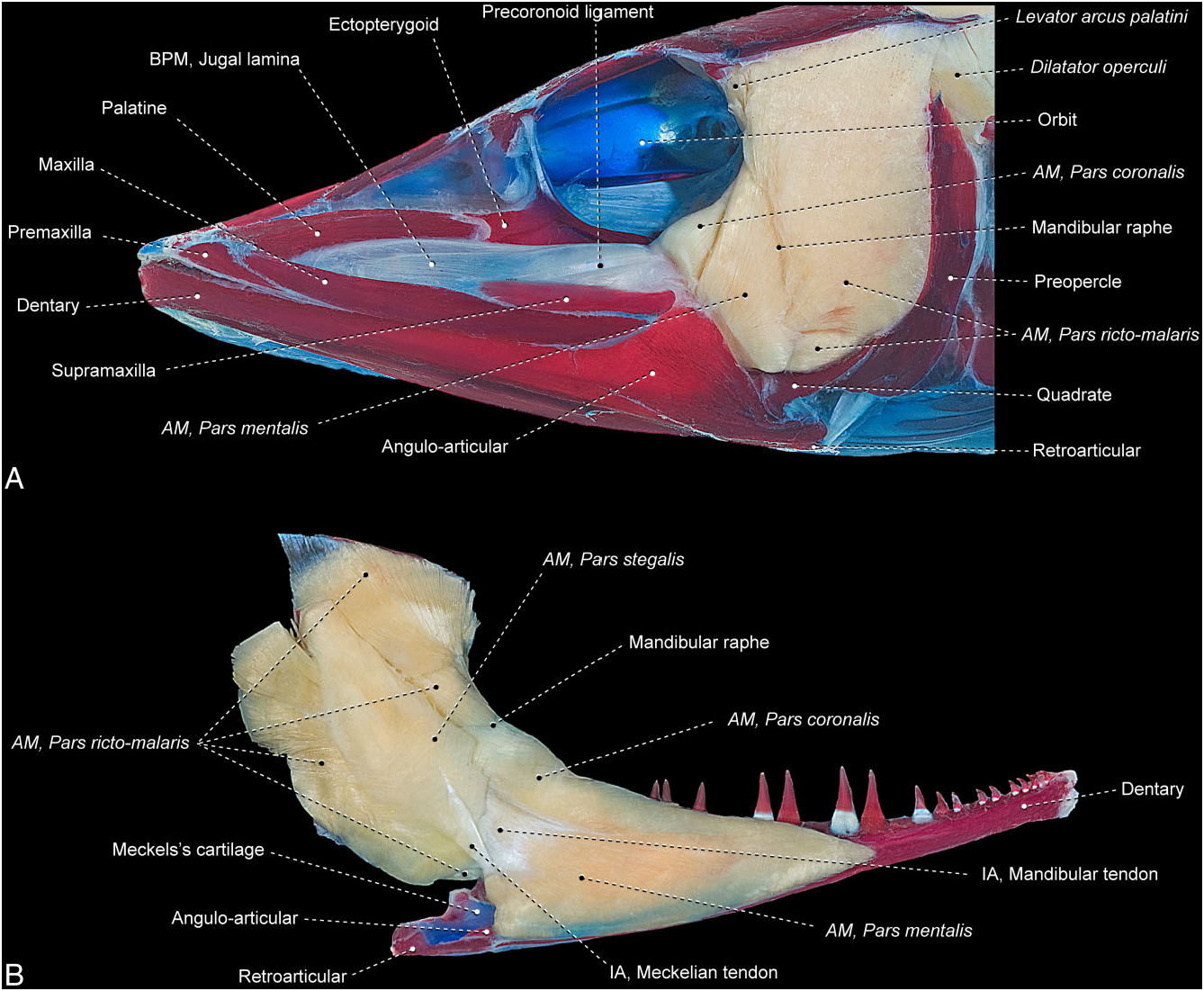
Esox americanus ( Fig. 24 View Figure 24 )
The segmentum facialis is divided into completely separate ricto-malaris and stegalis sections. The rictomalaris has an intricate architecture consisting of some differentiated, but not separated, bundles of muscle fibres. Notwithstanding this differentiation, none of these recognizable bundles seems to fully correspond to either the rictalis or the malaris. The combined rictomalaris originates from the quadrate, symplectic, preopercle, and hyomandibula. An inner group of fibres from this section passes through an ovoid fenestra bordered by the preopercle and hyomandibula to attach to a prominent posterodorsal process of the hyomandibula that contacts the opercle. The rictomalaris inserts medially on the mandibular tendon and laterally on the mandibular raphe. As in Umbra , a welldifferentiated precoronoid ligament is present, but this tissue band does not serve as the attachment site for any muscle fibres.
The stegalis originates primarily from the metapterygoid, with a few fibres arising from the posterodorsal process of the hyomandibula and passing through the aforementioned fenestra delimited by the hyomandibula and preopercle. The section inserts on the coronomeckelian via the meckelian tendon.
The ramus mandibularis trigeminus nerve could not be observed in the examined material.
The segmentum mandibularis is a massive element distinctly subdivided posterolaterally into the coronalis and mentalis sections. Both sections originate from the broad mandibular raphe and the medial face of the mandibular tendon, with a few posteromedial fibres arising from the meckelian tendon. The lateral-most fibres of the mentalis insert along the posterodorsal margin of the coronoid process of the angulo-articular. Towards their insertions, the coronalis and mentalis become continuous with one another and insert jointly on the medial surfaces of the angulo-articular, dentary, and Meckel’s cartilage.
Remarks
To facilitate the discussion, we follow the traditional classification under which the Esocoidei is divided into two families, the Esocidae and the Umbridae ( Greenwood et al., 1966; Nelson, 1972; Eschmeyer & Fong, 2013). Morphology of the adductor mandibulae is relatively consistent across the Umbridae . In all genera of this family the segmentum mandibularis extends beyond the posterodorsal margin of the lower jaw and the rictalis and malaris are differentiated from each other only along their anterolateral portions where they assume a bipinnate arrangement ( Fig. 23 View Figure 23 ; Williams, 1987, 1997). In most umbrids, the entire segmentum facialis converges to the intersegmental aponeurosis and lies internal to the ramus mandibularis trigeminus nerve. In Umbra limi and Umbra pygmaea the insertion of the ricto-malaris shifts laterally, with this section passing lateral to the ramus mandibularis trigeminus nerve and attaching tendinously to the lateral surface of the lower jaw ( Fig. 23A View Figure 23 ; Williams, 1987, 1997). Williams (1987, 1997) suggested that in Dallia , the muscle section lying anteromedial to the main facial muscle mass (his A2.dm?) could correspond to a part of the segmentum mandibularis (his Aw) that expanded posteriorly to such an extent that it partially attaches to the suspensorium. Comparisons across the Esocoidei, which also has part of the segmentum facialis greatly expanded posteriorly (see Figs 23 View Figure 23 , 24 View Figure 24 ), strongly support this assumption as the most parsimonious hypothesis of homology for this muscle division.
Multiple prior studies have discussed the adductor mandibulae of Esox , the sole extant genus of the Esocidae ( Vetter, 1878; Adams, 1919; Takahasi, 1925; Eaton, 1935; Van Dobben, 1935; Winterbottom, 1974; Williams, 1987, 1997; Sanford, 2000). The morphology of this muscle is relatively similar across all reported species of Esox . Contrary to the morphology of umbrids, esocids have the ricto-malaris (the A2 of most previous authors) well separated from the stegalis (the A3 of most previous authors) and the ricto-malaris originates from both the suspensorium and the neurocranium. Adams (1919) adopted a less common terminology, but as his illustrations are derived from those of Vetter (1878), we can confidently infer the homologies of the muscle divisions discussed by Adams (see Synonymy). Most previous studies of several species of Esox reported a ricto-malaris not differentiated into subsections and these observations agree with the condition in the examined sample of Esox americanus . Williams (1987, 1997) and Winterbottom (1974), alternatively, reported a posteri- or partial separation between a presumed malaris arising from the neurocranium and a rictalis with an origin on the suspensorium in Esox lucius and Esox americanus , respectively.
The segmentum mandibularis in esocids is posteriorly expanded to such an extent that some authors misidentified some of its parts as being components of the segmentum facialis. As Winterbottom (1974) conclud- ed and we concur, the sections in Esox termed the A 3 β by Vetter (1878) and A 1 β by Takahasi (1925) actually correspond to a dorsal division of Winterbottom’s (1974) Aw (= segmentum mandibularis), i.e. the coronalis.
Synonymy
Segmentum facialis
A 2: Van Dobben (1935): Esox .
A2: Williams (1987): Dallia , Novumbra , Umbra ; Williams (1997): Dallia , Umbra .
A 2 A 3: Takahasi (1925): Esox .
A2A3: Winterbottom (1974): Esox , Umbra .
Pars ricto-malaris
A 1 A 2: Eaton (1935): Esox , Umbra .
A 2: Takahasi (1925): Esox ; Vetter (1878): Esox .
A2: Sanford (2000): Esox ; Williams (1987): Esox ; Williams (1997): Esox ; Winterbottom (1974): Esox .
Adm 1: Adams (1919): Esox .
Pars rictalis
A2β: Williams (1987): Esox ; Williams (1997): Esox ; Winterbottom (1974): Esox .
Pars malaris
A2α: Williams (1987): Esox ; Williams (1997): Esox ; Winterbottom (1974): Esox .
Pars stegalis
A 3: Eaton (1935): Esox , Umbra ; Takahasi (1925): Esox ; Vetter (1878): Esox .
A3: Sanford (2000): Esox ; Williams (1987): Esox ; Williams (1997): Esox ; Winterbottom (1974): Esox .
Adm 2 plus Adm 3: Adams (1919): Esox .
Segmentum mandibularis
A2.dm? [ sic]: Williams (1987): Dallia ; Williams (1997): Dallia .
Adm 4: Adams (1919): Esox .
Aw: Sanford (2000): Esox ; Williams (1987): Esox , Novumbra , Umbra ; Williams (1997): Esox , Umbra ; Winterbottom (1974): Esox , Umbra .
Pars coronalis
A 1 β: Takahasi (1925): Esox .
A 3 β: Vetter (1878): Esox .
Pars mentalis Aω: Vetter (1878): Esox .
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.