
Nephrocerus Zetterstedt (1838)
|
publication ID |
https://doi.org/ 10.5281/zenodo.171339 |
|
DOI |
https://doi.org/10.5281/zenodo.6264623 |
|
persistent identifier |
https://treatment.plazi.org/id/03DC8797-FF95-FFCF-FEC3-F98BFC27F8CF |
|
treatment provided by |
Plazi |
|
scientific name |
Nephrocerus Zetterstedt (1838) |
| status |
|
Nephrocerus Zetterstedt (1838) View in CoL View at ENA
Nephrocerus Zetterstedt, 1838: 151 View in CoL . Type species: Nephrocerus lapponicus Zetterstedt, 1838 View in CoL , by monotypy.
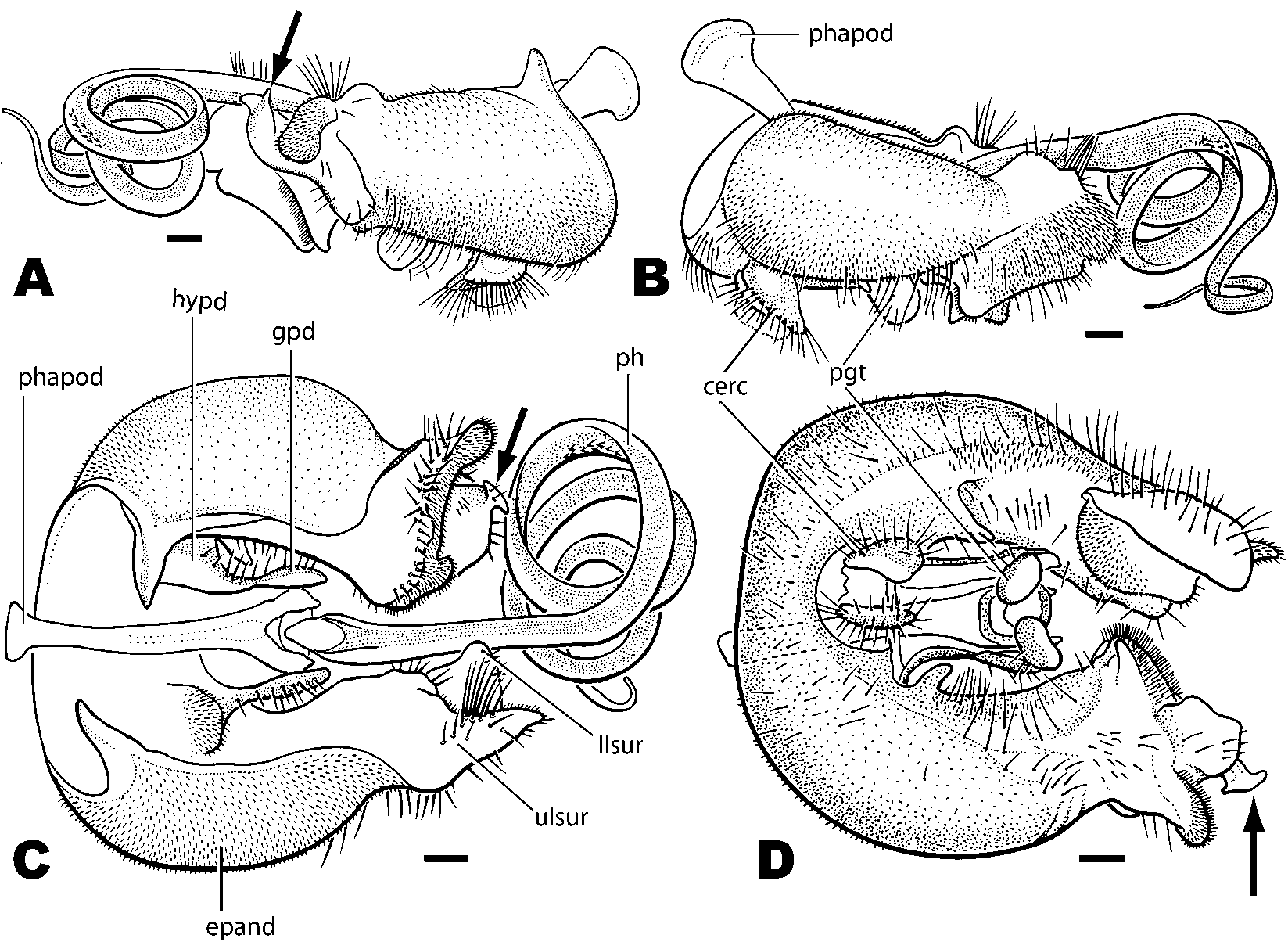
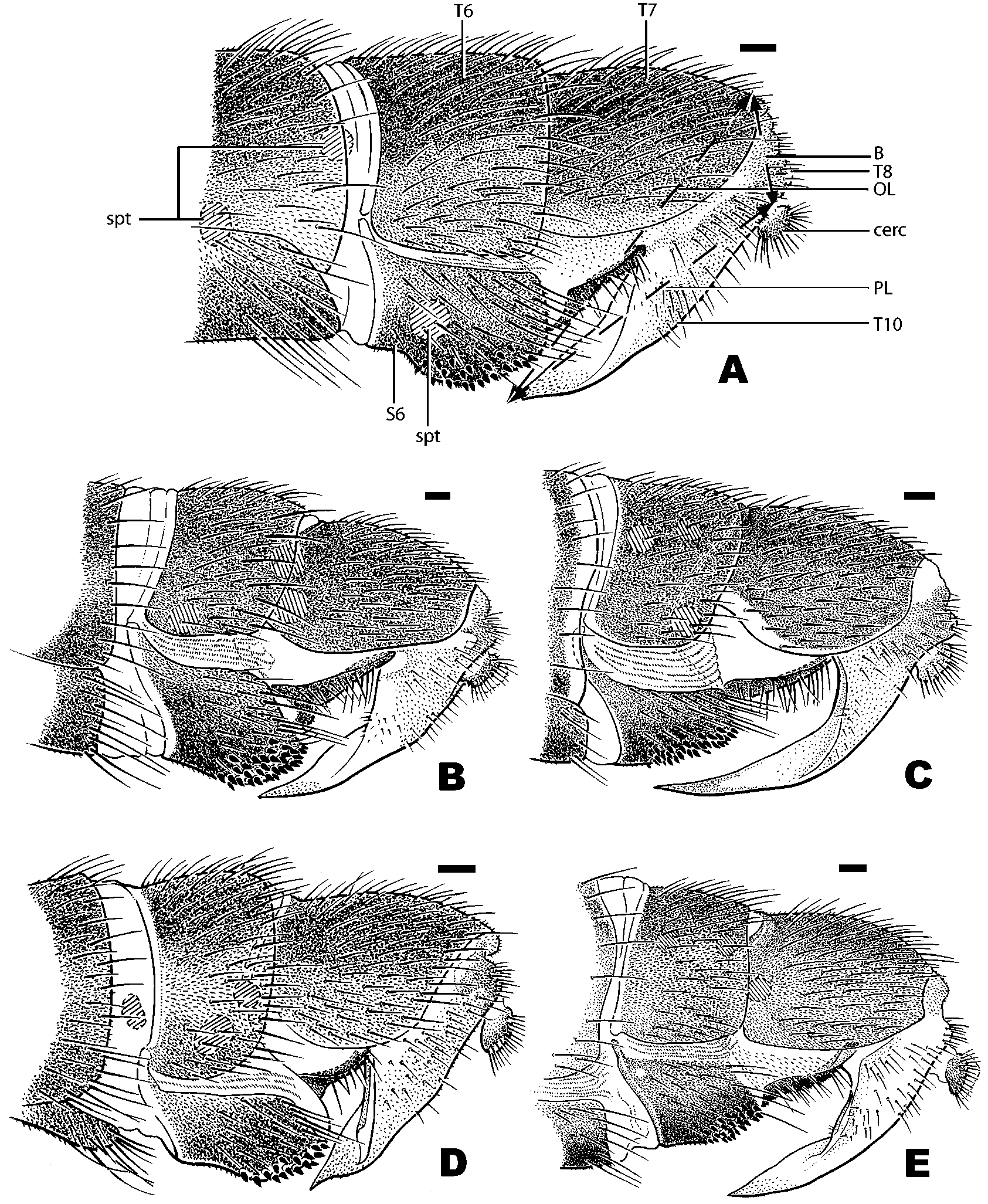
Description (adapted from Morakote (1988)): Male: Body length 5.7–12.0 mm. Head: hemispherical ( Fig. 1 View FIGURE 1 C); eyes touching on frons, hind margin of eyes deeply excised medially ( Fig. 1 View FIGURE 1 C); ocellar triangle bare, ocelli slightly raised ( Fig. 1 View FIGURE 1 B); frons and face densely silverpubescent, pubescence longer and denser on face; 1st flagellomere largest antennal segment, reniform ( Fig. 1 View FIGURE 1 C); anterior margin of scape and pedicel with many bristles that are slightly longer on pedicel; occiput moderately wide, sparsely covered with short hairs ( Fig. 1 View FIGURE 1 A). Thorax: proepisternum with a fan of bristles; postpronotal lobe enlarged, protruding ( Fig. 1 View FIGURE 1 A); 1–2 notopleural setae, 1–2 supraalar seta; dorsocentrals increasing in size posteriorly and terminating in 1–4 strong setae; 1–4 setae on postalar callus; scutellum with 2–8 long marginal bristles. Legs: long and slender, with dense pubescence and bristles; first and second tarsal segments very long, tarsi longer than femora and tibiae. Wings: 5.6–12.0 mm; long, about three times as long as broad; pterostigma absent; RM situated in basal third to fourth of discal medial cell; M2 present, long but never reaching wing margin. Abdomen: long and narrow, densely covered with pubescence and bristles; tergite 2 longest; tergites 7 and 8 twisted to the left; tergite 7 scarcely visible from above; syntergosternite 8 large, swollen. Genitalia: epandrium enlarged, horseshoeshaped; surstyli asymmetrical; hypandrium reduced to small plate, articulated laterally with basal part of gonopod ( Fig. 3 View FIGURE 3 C); postgonites situated at ventral base of phallus, small, oval ( Fig. 3 View FIGURE 3 D); phallus coiled, long when extended ( Fig. 3 View FIGURE 3 C); phallapodeme welldeveloped ( Fig. 3 View FIGURE 3 C). Female: As male except: eyes narrowly dichoptic; sternite 6 with numerous, short, tacklike spines on posterior half ( Fig. 4 View FIGURE 4 A); tergite 7 relatively long; ovipositor with very short base and short bladelike piercer.
Overview
Nephrocerus are large, yellow and black pipunculids that are more likely to be mistaken for flower flies ( Syrphidae ) than for other pipunculids. Head shape, ovipositor structure, wing venation (including the lack of a spurious vein) and general gestalt allow them to be differentiated from syrphids ( Fig. 1 View FIGURE 1 C). They can be easily diagnosed from other pipunculids by the shape of the eyes and antennae (see above). Protonephrocerus ( Chile only) is the only other pipunculid genus with the hind margin of the eye excised but they have the first flagellomere pointed ventrally (reniform in Nephrocerus ). A key to the world genera of Pipunculidae is available in Skevington & Yeates (2001).
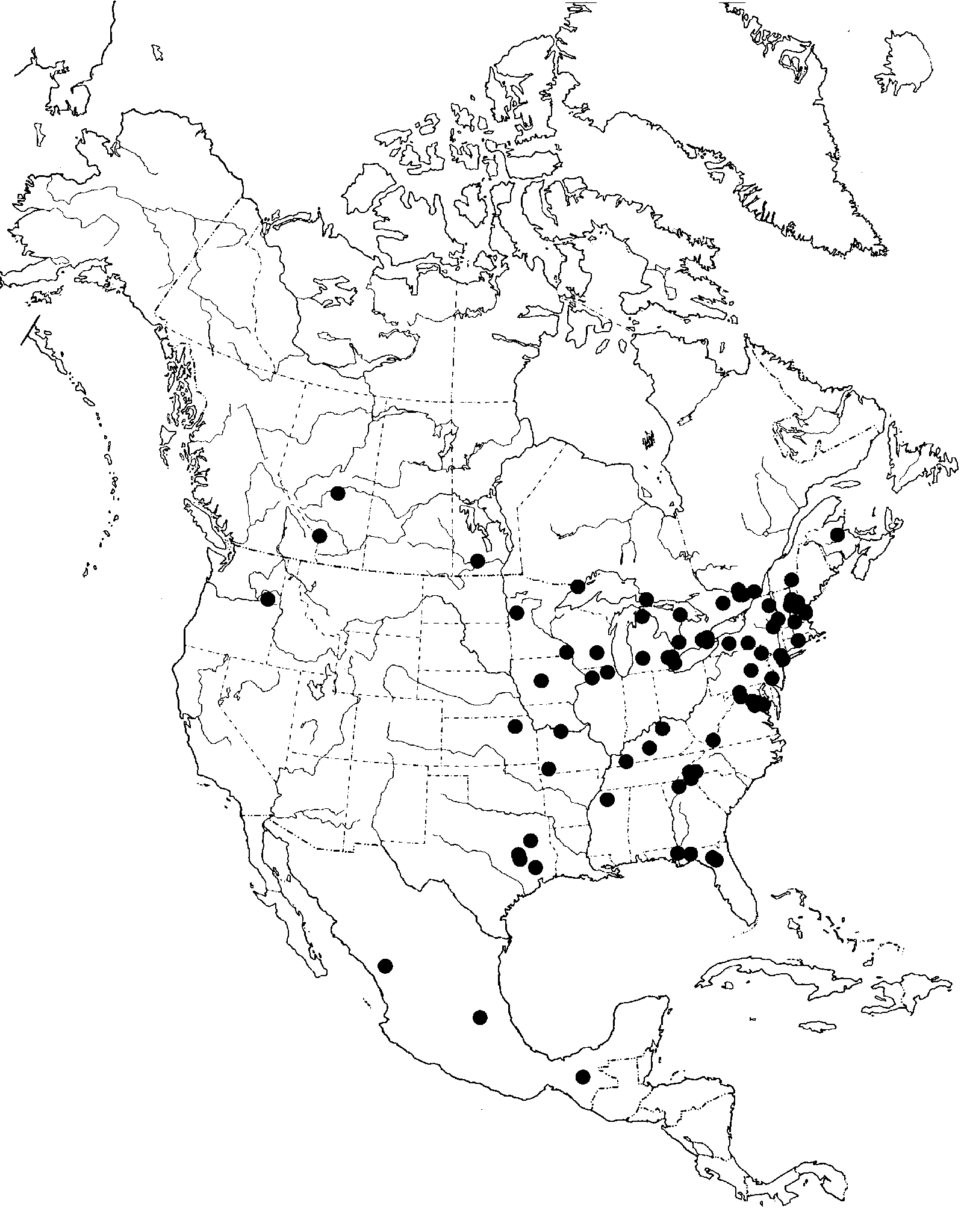
Nephrocerus is a northern temperate genus, previously documented only from the Palaearctic and Nearctic regions. The distribution of all New World specimens, including unidentified specimens, is shown in Figure 14 View FIGURE 14 . The New World distribution is distinctive. Only three specimens representing two species have been collected west of 100º W (basically a line running through western Manitoba south through western Kansas and the middle of Texas) in the USA and Canada. Three additional female specimens have been collected in Mexico, one as far south as Chiapas (16º45' N) ( Fig. 14 View FIGURE 14 ). This extends the known range of the genus into the Neotropical Region and suggests that we might expect to find Nephrocerus south into the mountains of Central America and in the West Indies.
Although the European species of Nephrocerus were revised in 1986 ( Grootaert & De Meyer 1986), new work in both Europe and Asia means that no single key will work for all Palaearctic species. The dominant three European species can still be identified using Grootaert & De Meyer (1986) ( N. flavicornis Zetterstedt , N. lapponicus Zetterstedt , and N. scutellatus Macquart ). However, identifications of these taxa should be checked against descriptions of species from Azerbaijan ( N. zaitzevi Kuznetzov (1990)) ; Russia ( N. fatalis Churkin (1991) and N. nevskajae Churkin (1991)) (to download an English translation of this paper, go to http://www.canacoll.org/ Diptera /Staff/Skevington/ Pipunculidae / Nephrocerus .htm); China ( N. auritus Xu & Yang (1997)) ; North Korea ( N. paektusanensis Kozánek & Kwon (1992)) ; and Japan ( N. flexus Morakote (1988) , N. grandis Morakote (1988) , N. japonicus Morakote (1988) , and N. spineus Morakote (1988)) .
Molecular and morphological evidence suggest that Nephrocerus occupies a key phylogenetic position within Pipunculidae between the ancestral Chalarinae and the diverse derived subfamily Pipunculinae ( Aczél 1948; Rafael & De Meyer 1992; Skevington & Yeates 2000). Only the relationship of Protonephrocerus and Nephrocerus is really disputed. The former appears to either be the sister genus to Nephrocerus ( Rafael & De Meyer 1992) or the sister of the Pipunculinae ( Aczél 1948; Skevington & Yeates 2000). Current treatment places Nephrocerus and Protonephrocerus as sister taxa in the Nephrocerinae . Resolving this relationship and discovering the hosts for these two genera will help us to understand the evolution and diversification of the family.
A species level phylogeny of Nephrocerus has never been attempted. Although there are good autapomorphic species level characters, there are few obvious synapomorphies to link groups of species. My efforts at creating a morphological phylogeny for all species of Nephrocerus met with little success. Unpublished molecular data (for Cytochrome c oxidase I subunit I COI) collected for another project generally supports the presence of two main lineages within the Nearctic taxa (the daeckei group and the slossonae group) (Skevington, unpublished data). These groups are easy to recognize based on the predominant colour of bristles on their bodies. The daeckei group has black hairs on the legs and abdomen while the slossonae group has yellow hairs. It is unclear whether or not these species groups will hold up on a World scale. Any attempts at producing a meaningful phylogeny for Nephrocerus will almost certainly have to rely on molecular data. Because most of the taxa are rare, collecting fresh material into alcohol for molecular work will be a challenge. Data from COI suggest that the Nearctic species are a cluster of recently diverging species. This gene did not exhibit enough sequence diversity to explore species limits or phylogenetics of the genus (Skevington, unpublished data). More rapidly evolving genes or gene regions such as ITS (internal transcribed spacer) and parts of the genes coding for rudimentary ( CAD) will be needed to study the phylogeny of Nephrocerus . Producing a cladistic hypothesis of relationships for Nephrocerus is clearly an important next step to research on this group of flies. This phylogeny will be useful for interpreting the interesting biogeographical pattern (essentially restricted to the Palaearctic and Eastern Nearctic Regions) exhibited by this genus. It will also serve as an important tool for understanding hostparasitoid relationships as more of these flies are reared.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Nephrocerus Zetterstedt (1838)
| Skevington, Jeffrey H. 2005 |
Nephrocerus
| Zetterstedt 1838: 151 |