
Asphalidesmus golovatchi Mesibov, 2009
|
publication ID |
https://doi.org/ 10.3897/zookeys.7.111 |
|
publication LSID |
lsid:zoobank.org:pub:2C6BD020-B54A-4119-9693-3231C9FCEFA6 |
|
DOI |
https://doi.org/10.5281/zenodo.3792456 |
|
persistent identifier |
https://treatment.plazi.org/id/D6CA5C87-8414-44C3-979A-080889549785 |
|
taxon LSID |
lsid:zoobank.org:act:D6CA5C87-8414-44C3-979A-080889549785 |
|
treatment provided by |
Plazi |
|
scientific name |
Asphalidesmus golovatchi Mesibov |
| status |
sp. nov. |
Asphalidesmus golovatchi Mesibov View in CoL , sp. n.
urn:lsid:zoobank.org:act:D6CA5C87-8414-44C3-979A-080889549785
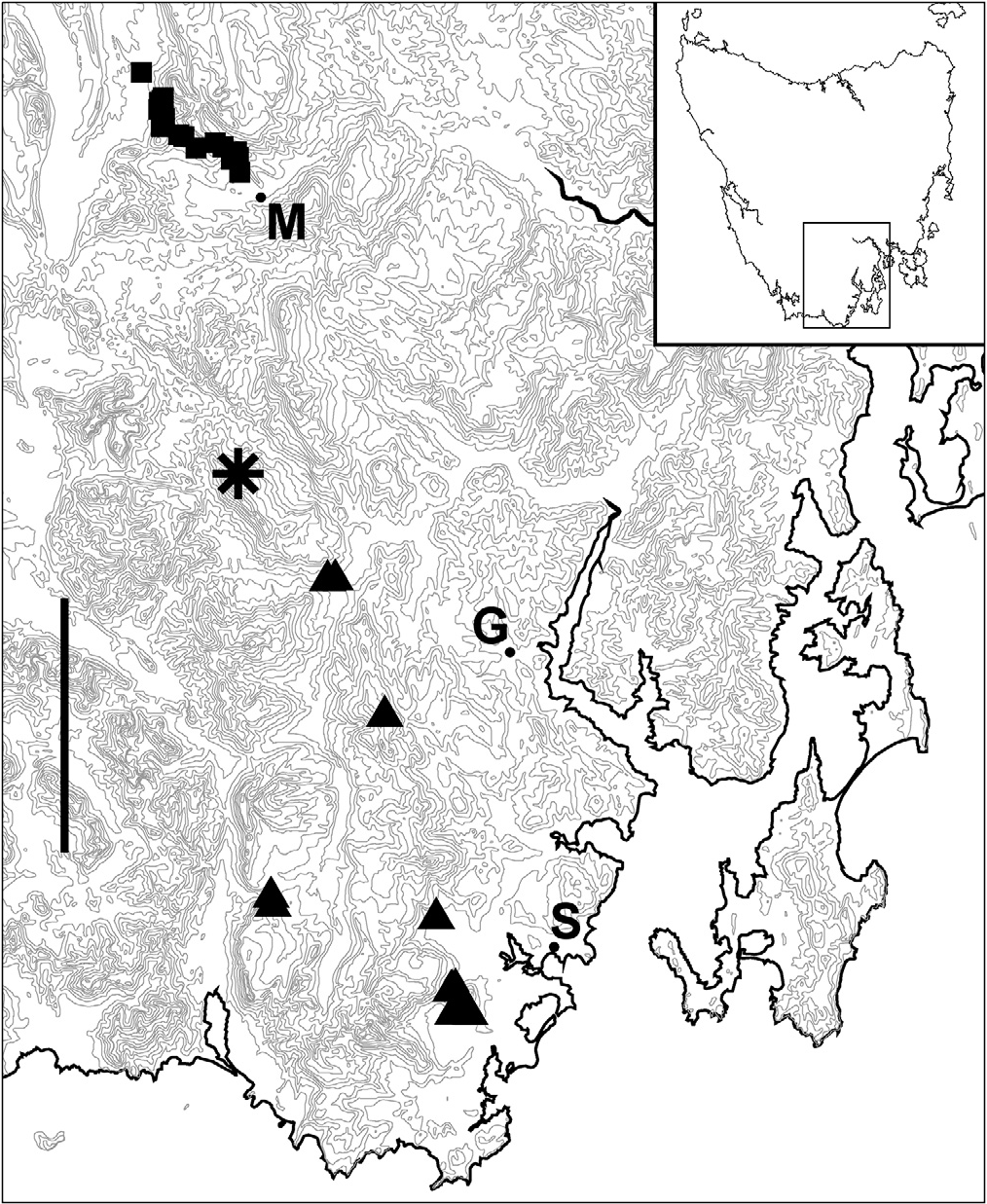
Figs. 1C, 1D View Figure 1 , 2C View Figure 2 , 6B View Figure 6 , 6C View Figure 6 , 7A View Figure 7 , 7B View Figure 7 , 8A View Figure 8 , 8B View Figure 8 , map Fig. 9 View Figure 9 .
Holotype. Male. Australia, Tasmania, Lake Osborne track, 43º13’04”S 146º46’03”E ± 100 m, 880 m, 7 February 2004, K. Bonham, QVM 23 View Materials :25723. GoogleMaps
Paratypes. 2 females, 2 stadium VI females, Resurgence Cave , Vanishing Falls karst, 43º23’S 146º38’E ± 1 km, 25 April 1992, S. Eberhard, QVM 23 View Materials :12956 GoogleMaps ; 2 males, 2 females, 1 stadium VI male, 2 stadium VI females, same details but 28 August 1992, S. Eberhard and V. Wong , QVM 23 View Materials :12949 ; 3 males, 3 females, 2 stadium VI females (includes male+female in copula), Spring Cave , Vanishing Falls karst, 43º23’S 146º38’E ± 1 km, 28 April 1992, S. Eberhard, QVM 23 View Materials :12971 GoogleMaps ; 1 male, Warra coupe WR001B, 43º05’48”S 146º41’55”E ± 100 m, 90 m, pitfall 254 emptied 14 April 2000, R. Bashford, QVM 23 View Materials :45660 GoogleMaps ; 1 stadium VI female, Mystery Creek Cave track, 43º27’39”S 146º51’11”E ± 100 m, 160 m, K. Bonham and R. and J. Francis, 25 February 2001, QVM 23 View Materials :24747 GoogleMaps ; 1 male, ca 100 m uphill from Mystery Creek cave , 43º27’42”S 146º50’57”E ± 100 m, 5 February 2006, K. Bonham, QVM 23 View Materials :46402 GoogleMaps .
Material examined. 4 females, 1 stadium VI female, Entrance Cave , Ida Bay karst, 43º28’S 146º51’E ± 1 km, 20 January 1985, S. Eberhard, sample IB10-9 GoogleMaps ; ‘ small sp. on mud, upper levels above final siphon’, QVM 23 View Materials :41576 ; 3 males, 3 females, 2 stadium VI males, Milk Run cave , Ida Bay karst, 43º29’S 146º51’E ± 1 km, 22 August 1985, S. Eberhard, sample IB38-4, ‘above stream at bottom - deep’, QVM 23 View Materials :12167 GoogleMaps ; 2 females, 2 stadium VI females, 1 stadium V female, Spider Den cave (NL-3), North Lune karst, 43º24’S 146º50’E ± 1 km, 5 February 1988, A. Clarke, QVM 23 View Materials :11671 GoogleMaps ; 1 stadium VI male, same details but 31 October 1988, sample 1088-15, QVM 23 View Materials :46558 ; 1 stadium VI female, same details but sample 1088-23, ‘under detritus at base of cave dark zone’, QVM 23 View Materials :46559 ; 1 stadium V female, Midnight Hole cave , Ida Bay karst, 43º28’S 146º51’E ± 1 km, 2 April 1989, S. Eberhard, sample IB11-4, QVM 23 View Materials : 12074 GoogleMaps ; 2 females, Ida Bay cave 46, 43º29’S 146º52’E ± 1 km, 23 March 1990, S. Eberhard, P1 chamber, QVM 23 View Materials :46561 GoogleMaps ; 1 stadium VI female, Huon River (Manuka Road), 43º05’46”S 146º42’28”E ± 100 m, 100 m, 30 April 1997, R. Mesibov, plot 1M1, QVM 23 View Materials :41564 GoogleMaps ; 1 stadium VI female, same details but plot 1M2, QVM 23 View Materials :46562 .
Diagnosis. Differs from A. leae and A. parvus in having long, tapering rather than short, convex paranota, and in ozopore opening well away from base of paranotum (at about midheight on body in lateral view) rather than just above paranotal base.
Description. Juveniles and cave-dwelling adults unpigmented, but as in Asphalidesmus leae and A. parvus , tergites of surface-dwelling adults are partly encrusted with soil particles and stained light yellow-brown. Surface-dwelling males ca 7 mm long, maximum diameter 0.7 mm, maximum width across paranota 1.2 mm.
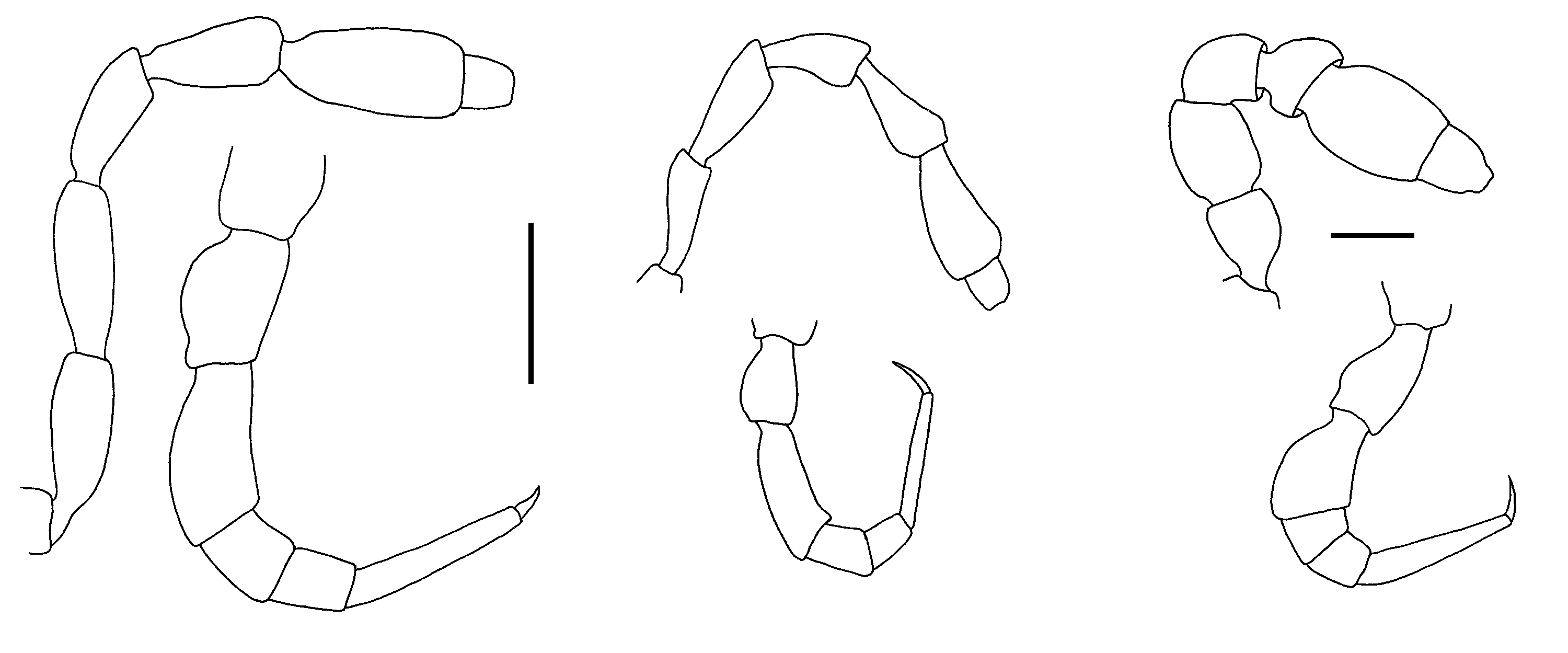
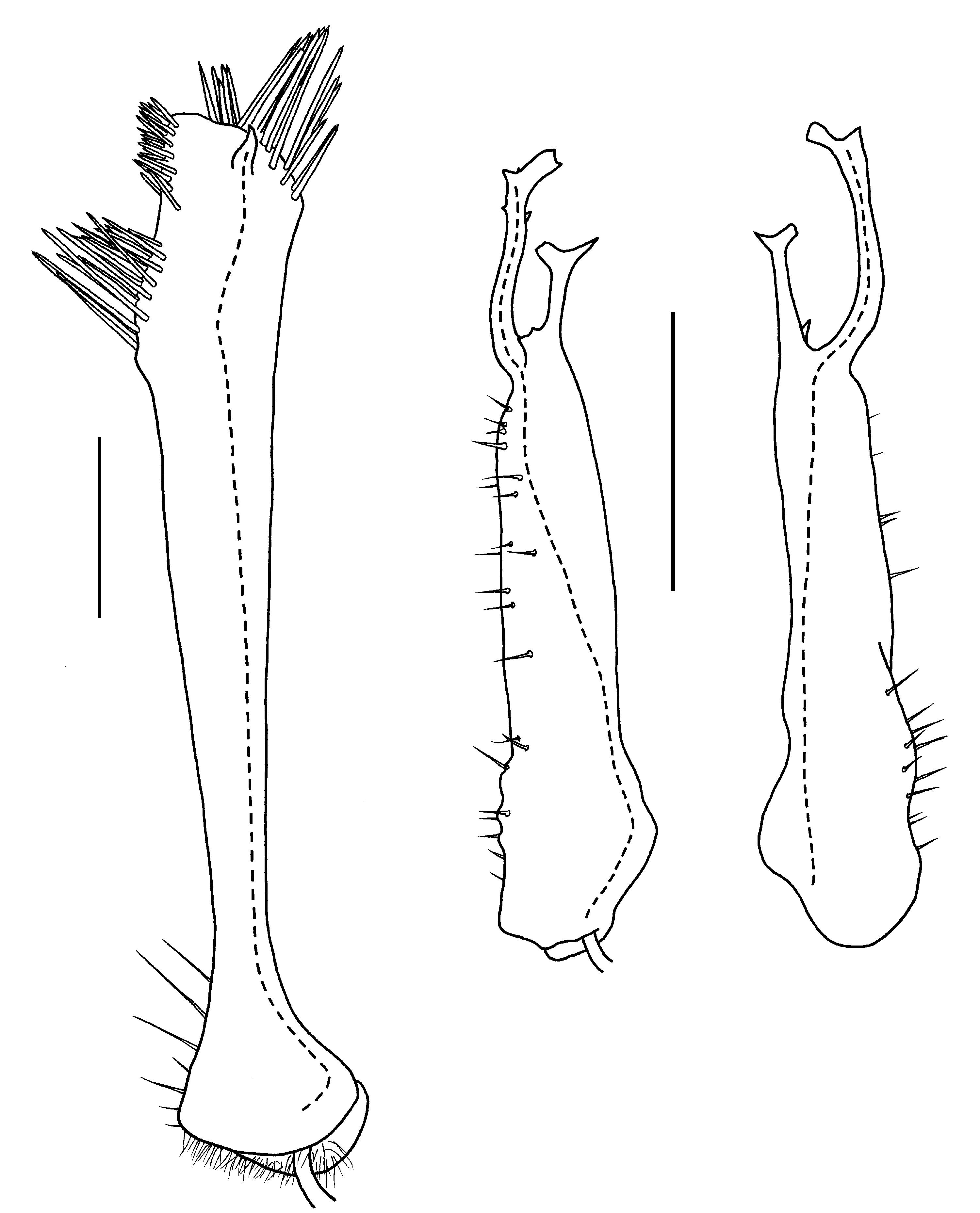
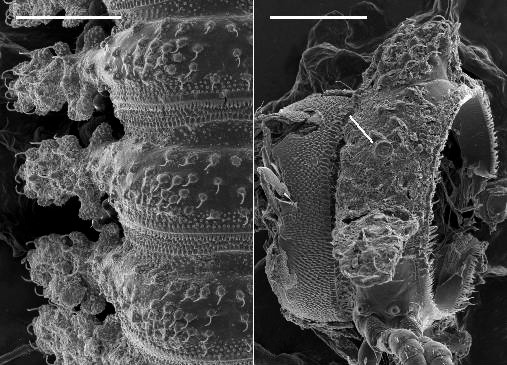
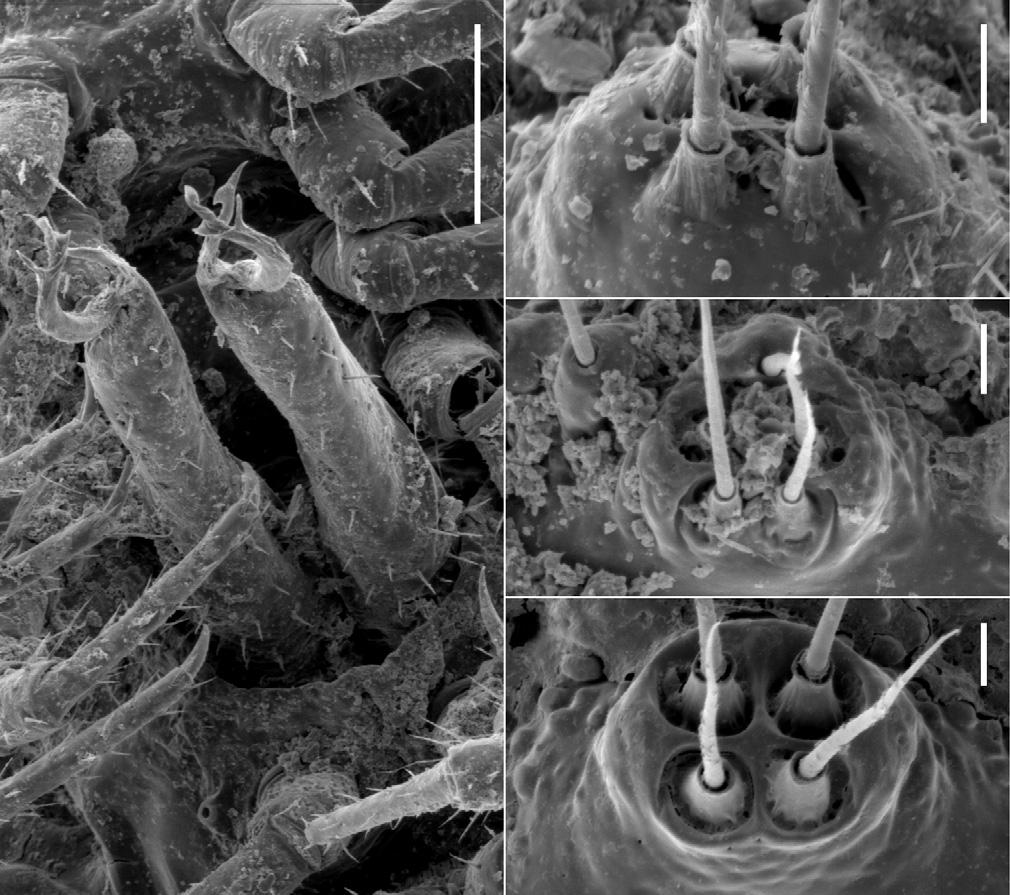
Male ( Fig. 1C View Figure 1 ) with head and last ring strongly flexed to face substrate. Head sparsely setose, as wide as collum; antennal sockets strongly impressed ventrolaterally, separated by about 1.5X socket diameter. Antenna ( Fig. 2C View Figure 2 ) short and thick; antennomere 6 widest and longest; relative antennomere lengths 6>>3>2>(4,5). Collum with anterior edge nearly straight, posterior edge broadly convex, corners blunt. Overall ring widths diminishing gradually from ring 3 posteriorly. Collum, metatergites and paranota with transverse zone of small tubercles ( Figs 7A, 7B View Figure 7 ), each bearing a stout, pointed seta; metazonites also with much smaller, non-setiferous tubercles anteriorly and posteriorly; prozonites with narrow band of longitudinal ridges just anterior to suture, elsewhere uniformly covered with very small protuberances with blunt, rounded tips directed slightly posteriorly. Limbus composed of long tabs with multi-toothed tips and a narrow, outwardly curving medial section. Tergites of rings 2-16 with paired, paramedian, dorsal projections ( Fig. 1D View Figure 1 ), each projection thick, rounded and directed slightly posteriorly; tergites of rings 17 and 18 each with one mid-dorsal, thick, rounded and posteriorly directed projection (dorsal projections absent in juveniles). Paranota of ring 2 greatly expanded laterally and anteriorly and strongly depressed, in lateral view masking head, antennae and collum. Paranota of rings 3-18 set low on body ( Fig. 1D View Figure 1 ), more or less hastate, strongly depressed; lateral extent diminishing gradually from anterior to posterior; anterior and posterior margins scalloped with small, usually discrete, rounded tabs. Ring 3 paranotum slightly overlapping ring 2 paranotum. Pore formula 5, 7, 9, 10, 12, 13, 15-18; ozopore very small, opening in slight depression in low, cylindrical structure well above base of paranotum ( Fig. 7B View Figure 7 ). Sternites somewhat longer than wide, longitudinal and transverse impressions well-defined. Pre-anal ring with a few dorsal marginal setae, epiproct not developed, hypoproct paraboloid. Spinnerets in square array ( Fig. 8B View Figure 8 ) in slight depression with low partition wall between dorsal and ventral pairs; sheath separated from seta by annular gap. Anterior legs ( Fig. 2C View Figure 2 ) short; prefemur and femur a little expanded dorsally; relative podomere lengths tarsus>(prefemur, femur)>(postfemur, tibia). No sphaerotrichomes or brush setae; very small tuberculation of metazonite surface extending onto coxa/trochanter. Spiracles ( Fig. 7B View Figure 7 ) opening on short, wide-rimmed elevations; on diplosegments with anterior spiracle above anterior leg and oriented anterolaterally, and posterior spiracle above and about midway between anterior and posterior legbases. Gonopore small, opening on distomedial projection of leg 2 coxa. Legpair 6, 7 bases well-separated to accommodate retracted gonopods, legpair 4, 5 bases a little less separated.
Gonopod aperture ovoid, about one-third prozonite width; posterolateral margins slightly raised. Gonocoxae small, truncated conical, weakly joined distomedially, with a few setae on basomedial and distolateral surfaces. Cannula prominent, inserting in shallow depression on basal surface. Telopodites separate, reaching to legpair 4 bases when retracted. Telopodite ( Figs 6B, 6C View Figure 6 , 8A View Figure 8 ) cylindrical, slightly tapering distally, with two slender branches arising at slight constriction at about three-quarters telopodite height; anteromedial branch bending laterally and terminating in ‘fishtail’ fork at about two-thirds height of posterolateral branch; posterolateral branch (= solenomere) curving posterolaterally, then slightly anteromedially before flattening and curling apically with two upright finger-like processes arising from curled, flat tip: a narrower, shorter, more lateral process, and a stouter, taller, more medial process bearing the end of the prostatic groove. Telopodite sparsely setose posterolaterally from near base to distal constriction. Prostatic groove running slightly anteriorly from insertion before running distally to telopodite constriction, then following curve of solenomere to tip.
Female as large as male; posterior margin of epigynum slightly raised medially; cyphopods not examined.
Distribution and habitat. Known from wet eucalypt forest and caves over ca 600 km 2 in far southern Tasmania from ca 100 m to 900 m ( Fig. 9 View Figure 9 ). Uncommon in forest. Sympatric over the whole of its range with A. parvus , which also occurs in caves.
Etymology. Adjective, genitive singular, for Sergei I. Golovatch, Russian diplodologist, who has generously given me advice on diplopodological problems and who has taken a particular interest in Asphalidesmus .
Remarks. Cave specimens of A. golovatchi are a little smaller than surface-dwelling specimens in the same stadium. In some of the mature cave specimens the paramedian dorsal projections are greatly reduced, although the median dorsal projections on the last two leg-bearing rings can be clearly seen.
The pair in copula from Spring Cave (QVM 23:12971) are in the usual position for mating Polydesmida . Although the male’s head is flexed strongly down, there is still a gap between it and the female’s head.
The spinnerets in A. leae are arranged as in A. golovatchi , while in A. parvus low partition walls divide the depression housing the spinnerets into four separate compartments ( Figs 8 View Figure 8 B-8D).

| VI |
Mykotektet, National Veterinary Institute |
| V |
Royal British Columbia Museum - Herbarium |
| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |