
Diprotodon, Owen, 1838
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2008.00387.x |
|
DOI |
https://doi.org/10.5281/zenodo.10546325 |
|
persistent identifier |
https://treatment.plazi.org/id/03DC87E5-D142-FFA8-2234-FE46FF0DFC6E |
|
treatment provided by |
Felipe |
|
scientific name |
Diprotodon |
| status |
|
DARLING DOWNS DIPROTODON
Molnar & Kurz’s (1997) audit of the Queensland Museum Darling Downs collections suggested that the large-form Diprotodon is eight times more common than the small form. However, that audit did not challenge the identifications previously placed on specimens, and the ratio could be inaccurate. The present data suggest that the small form is represented by at least 30 individuals and the large form by 21 individuals. The differing results demonstrate the high degree of morphological similarity between the large- and small-form Diprotodon , and highlights the difficulties in assigning isolated specimens to a specific size class.
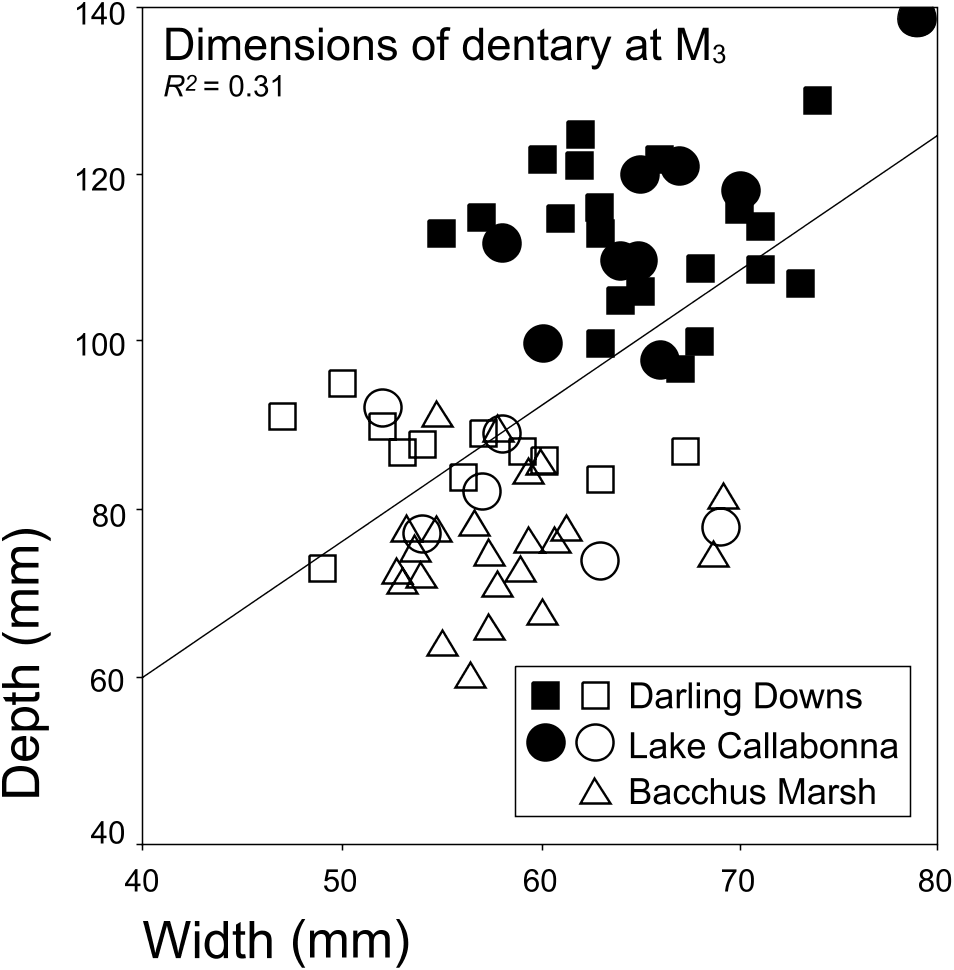
Teeth of Darling Downs Diprotodon large-form individuals are slightly larger than corresponding teeth of small-form individuals. A similar morphometric relationship is exhibited in sexually dimorphic extant grey kangaroo ( Macropus giganteus Shaw, 1790 ) populations collected from the Warwick region (see supplementary Appendix S2 for list of specimens examined), southern Darling Downs ( Fig. 13 View Figure 13 ). In terms of dentary cheek teeth dimensions, grey kangaroo males are mostly ~4–10% larger than females, although a considerable degree of overlap exists between sexes. On average, COV and R 2 values indicate that the correlation between length and width of Darling Downs Diprotodon lower molar teeth ( Fig. 10 View Figure 10 ) is similar to values for the corresponding teeth of the modern grey kangaroo population ( Fig. 13 View Figure 13 ) from the Darling Downs ( Tables 3, 4). However, examination of COV values shows that Darling Downs Diprotodon upper molar teeth are slightly more variable than for corresponding teeth of modern populations of grey kangaroos.
Following criteria established in the Introduction, the data collected from the assemblage are consistent with the hypothesis that Darling Downs Diprotodon is represented by a single, sexually dimorphic species. That interpretation is supported by the following observations: (1) other than differences in the size and shape of the dentary, there are no significant or consistent dental morphological differences (particularly in the systematically important upper premolar) between large- and small-form Darling Downs Diprotodon ; (2) both teeth and dentary size are consistent with a bimodal body size distribution within the assemblage; and (3) both size classes are temporally and spatially coeval.
In sexually dimorphic mammals, males are generally larger than females ( Andersson, 1994). However, there are a few rare cases where the female is the larger sex (e.g. extant spotted hyena, Crocuta crocuta , although in that case, dimorphism is reflected in body size only and does not extend to tooth morphometrics) ( Van Horn, McElhinny & Holekamp, 2003). Within Darling Downs Diprotodon and other Diprotodon assemblages in general, it is not possible unambiguously to assign gender on the basis of morphometric or morphological differences in dentary and tooth size. However, by drawing analogy to all extant megaherbivores, the male is almost invariably larger than the female (although extant black rhinos, Diceros bicornis , are slightly more monomorphic) ( Owen-Smith, 1988). Additionally, the male is significantly larger than the female in all sexually dimorphic extant marsupials greater than 5 kg in mean sexed body weight (predominantly macropodoids) ( Jarman, 1989). By drawing such analogies for Diprotodon , it is most likely that the large form is male and the small form is female. However, that hypothesis remains difficult to test. Although placentals show morphological and morphometric skeletal differences reflective of parturition and thus of gender ( Averianov, 1996; Lister, 1996), such differences are less obvious in marsupials ( Tague, 2003). Some extant marsupials do show gender-related differences in skeletal limb portions ( Jarman, 1989); however, demonstrating similar post-cranial, gender-related osteological differences in Diprotodon is difficult because there are very few (if any) completely articulated or associated Diprotodon skeletons for comparison.
In most cases, COV values for Bacchus Marsh Diprotodon cheek teeth dimensions are more similar to those of the modern grey kangaroo population than to the Darling Downs Diprotodon assemblage ( Tables 2– 4). That may reflect: (1) a different temporal sampling range at Bacchus Marsh than at the Darling Downs (presumably of less duration for Bacchus Marsh); (2) a condition inherent to either the Bacchus Marsh or the Darling Downs Diprotodon assemblages; and/or (3) simply reduced variation due to one size class being represented. Testing such hypotheses is difficult due to the paucity of material available for study. However, an extended temporal period of sampling would support recent sedimentological and taphonomic interpretations regarding the attritional nature of the majority of Darling Downs fossil assemblages ( Price & Sobbe, 2005; Price & Webb, 2006). Additionally, taphonomic data for Bacchus Marsh indicate that the Diprotodon assemblage accumulated during a single, mass mortality event, suggesting that biotic sampling occurred over a particularly short temporal scale ( Long & Mackness, 1994).
At face value, the data compiled for Bacchus Marsh Diprotodon are not entirely consistent with an interpretation of sexual dimorphism within the assemblage. As with the Darling Downs, there are no consistent dental morphologies to warrant separation of more than one morphospecies in the assemblage. However, morphometric data suggest that the single morphospecies was monomorphic in body size, but similar in size to the small-form (?female) Darling Downs Diprotodon ( Fig. 8 View Figure 8 ; Tables 1–3). It would be expected that a sexually dimorphic assemblage would contain members of both sexes (i.e. both size classes). However, biological and taphonomic information provides some evidence that the assemblage could represent a gender-segregated herd. Extant sexually dimorphic megafauna commonly occur in gendersegregated herds ( Owen-Smith, 1988; Haynes, 1991; Berger et al., 2001). As the assemblage is thought to represent individuals trapped in muds of a drying marsh ( Long & Mackness, 1994), it is possible that a single herd was sampled. Long & Mackness (1994) suggested that the preponderance of subadult to adult individuals over juvenile and mature individuals in the Bacchus Marsh Diprotodon assemblage reflected a population that was stressed by drought. Similar demographic profiles have been observed in droughtstressed sexually dimorphic marsupials such as the extant red kangaroo, Macropus rufus Desmarest, 1817 , and the grey kangaroo ( Newsome, 1965; Kirkpatric & McEvoy, 1966). Red and grey kangaroo fecundity decreases in drought conditions and adults tend to be the dominant surviving group ( Newsome, 1965; Kirkpatric & McEvoy, 1966). Thus, the overabundance of small-form (?female) individuals in the Bacchus Marsh Diprotodon assemblage may be explained by drought effects. Norbury, Coulson & Walters (1988) observed that male-biased mortality is extenuated by drought conditions in sexually dimorphic marsupials such as the western grey kangaroo, Macropus fuliginosus Desmarest, 1817 . Such biased mortality patterns are similar in all age classes and reflect disparate energy costs imposed by sex differences in body size and mobility ( Norbury et al., 1988). Thus, at Bacchus Marsh, large-form (?male) Diprotodon individuals of all age classes may have a succumbed to drought conditions before small-form (?female) Diprotodon individuals, and hence were not sampled in this chance event where one herd died around a drying water course. The other assemblages that contain both large- and small-form individuals all represent broader attritional accumulation processes that would be unlikely to sample only a single herd.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |