
Microgomphus alani, Kosterin, Oleg E., 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4114.3.9 |
|
publication LSID |
lsid:zoobank.org:pub:7CC2B34B-139A-46A9-BD0E-C79E51F7E41F |
|
DOI |
https://doi.org/10.5281/zenodo.6087608 |
|
persistent identifier |
https://treatment.plazi.org/id/03DD6011-D523-5C13-FF14-FE1B3E7FFAC7 |
|
treatment provided by |
Plazi |
|
scientific name |
Microgomphus alani |
| status |
sp. nov. |
Microgomphus alani View in CoL sp. nov.
( Figs. 1–2 View FIGURE 1 View FIGURE 2 , 3 View FIGURE 3 a,c, 4)
Microgomphus sp. — Kosterin 2010: p. 64–65, fig. 43 below. Microgomphus sp. — Kosterin et al. 2012.
Holotype: ♂ (preserved in alcohol, Fig. 1 View FIGURE 1 a,c; 2a-f,h; 3a-b,e-f, 4), Cambodia, Koh Kong Province, 17 km ENE of Koh Kong, ‘Macromia Rivulet’, 11°40'17'' N, 103°07'28'' E, 296 m a.s.l., 3 vi 2014, O. Kosterin leg.; deposited in Naturalis Biodiversity Centre, Leiden, the Netherlands ( RMNH). Paratype: ♂ (appendages twisted; preserved dry, Fig. 1 View FIGURE 1 b, 2g, 3c-d), Cambodia, Koh Kong Province, 6.5 km SW Thma Bang village, the ‘ Microgomphus River’, the slow reach upstream from the bridge, 11°38'42'' N, 103°23'49'' E, 341 m a.s.l., 15 iv 2010 (in the author’s collection).
Etymology. The new species is named after Alan Andrews in recognition of his amity and aid: on the second day of our acquaintance he introduced me to the Cardamoms and showed me an excellent dragonfly habitat where we found the paratype.
Holotype male. Body blackish brown (because of teneral condition) with yellowish markings ( Fig. 1 View FIGURE 1 a).
Head ( Fig. 2 View FIGURE 2 b). Face black with pale pattern. Labium pale but apical parts of mentum and movable hooks brownish. Mandible bases pale. Labrum with a pair of large oval spots on either side. Anteclypeus with a broad subtriangular spot along upper margin, with apex extending to its centre. Lower lateral parts of postclypeus pale. Frons dorsal surface with a broad pale stripe narrowly interrupted at middle. Vertex and occiput dark. Occiput with two very slight waves, set with long hairs. Sparse shorter hairs scattered over face, more behind lateral ocelli, frons dorsal surface, lower margins of postclypeus and labrum.
Thorax. Prothorax anterior lobe yellow; sides of median lobe pale with indistinct border of that colour, the rest brown.
Synthorax ( Fig. 2 View FIGURE 2 a,b) with the following pale pattern. Dorsal stripes broad, fused to short, broad stripes of mesothoracic collar at their lower ends to form two inverted 7s; collar stripes not being fused to each other at medial carina. Barely noticeable traces of antealar spots (which most probably would disappear with maturity). A broad pale stripe occupies ca 2/3 of mesepimeron width, shifted towards its anterior margin so that it leaves dark borders, of which anterior is about twice as narrow as posterior. Another broad pale stripe occupies ca 2/3 of mesepisternum width but shifted to its posterior margin. Metepimeron pale except for a dark anterior border and anteroventral corner. Inframesepisternum with an elongate vertical pale spot in lower part; inframetepisternum pale but anteroventral corner dark. Generally the lateral synthoracic pattern consists of two dark parallel lateral stripes, along mesepimeron/metepisternum suture (this one includes spiracle) and metepisternum/metepimeron suture. Ventral side pale.
Coxae with pale vertical outer stripes. Legs brown with black spines, claws pale with black teeth and apices. Hind femora reach anterior apex of vesica spermalis.
Wings ( Fig. 1 View FIGURE 1 c) hyaline, basally with a yellowish tint gradually disappearing at ca halfway between base and nodus; venation brownish. Antenodals (1st and 5th primary) 12 (right)–13 (left) on forewings, 10 on hindwings; postnodals 9 on forewing (left missing), 9 (right)–7 (left) on hindwings. Crossveins between Arc and R1-R4 junction 6 above Rs and 5 (right)–6 (left) below Rs on fore wing, 3 (right)–4 (left) above Rs and 3 below Rs on hindwing. Cubitanal crossveins 2 on forewing, 1 on hindwing. Anal triangle almost equilateral, 3-celled. Tornus approximately rectangular but slightly rounded, wing margin between base and tornus moderately sinuous; membranula absent. Pterostigmata pale brownish, almost transparent; those on forewing cover 4 cells below, on hindwing 3.5 cells below; lower bordering vein more swollen than upper one, somewhat folded.
Abdomen brown with pale marking as follows ( Fig. 1 View FIGURE 1 a): S1 with dorsum occupied by a broad triangular spot, pale ventrally, with dark background forming narrow ventrad projections. S2 with a narrowly triangular dorsal spot directed caudally and pale ventrally, with dark background forming broad rounded ventrad projections behind auricles and dark border at segment posterior margin. S3–6 with fine dorsal pale streaks broadening at anterior margin and small ventrolateralpale spots. S7 with a broad, even anterior pale ring occupying about ¼ of its length. Rest of abdomen dark.
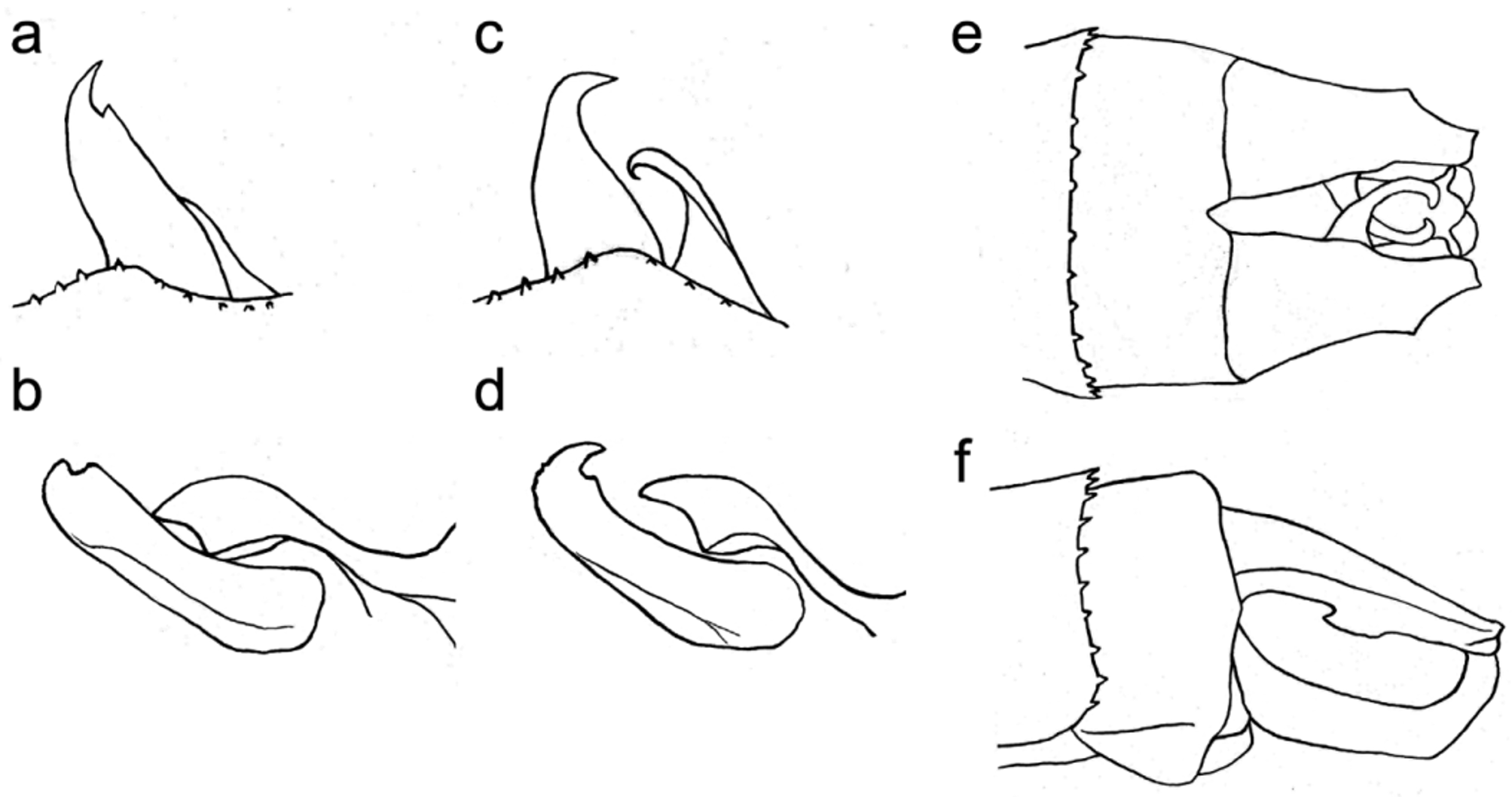
S10 posterior margin with a small but deep central triangular notch. Dorsal surface of cerci pale but distal half of outer margins brown (with indistinct border of colours) and apices black; the rest brown. Main branch of cerci flat, in dorsal view ( Fig. 2 View FIGURE 2 c, 3d) parallel, elongate triangular, with truncated, slightly sinuate apices and distinct blunt angulations at outer margins at 2/3 length. Inner arms of cerci arise just behind middle of inner margins, sickle-shaped, strongly curved and hooked, those of opposite cerci cross and strongly superimpose upon each other, not reaching apices of main branch of cercus. In lateral view ( Fig. 2 View FIGURE 2 d, 3f), inner arms only very slightly declining from main branch. In dorsal view ( Fig. 2 View FIGURE 2 c, 3d), epiproct protrudes slightly behind cerci and its two hooked, slightly divided, pointed apices visible. In lateral view ( Fig. 2 View FIGURE 2 d, 3f) almost straight but shortly before cercus tip bends up with apices hooked behind. In ventral ( Fig. 2 View FIGURE 2 e) and posteroventral view, its main body concave, upturned part divided but both arms run parallel in contact with each other until terminating in roundly diverging and pointed apices.
Accessory genitalia ( Fig. 2 View FIGURE 2 f) light brown, hirsute. In lateral view anterior hamuli hidden behind posterior hamuli ( Fig. 2 View FIGURE 2 f, 3a); in ventral view ( Fig. 2 View FIGURE 2 f, 3b) anterior hamulus has a narrow ‘neck’ and then expands to a broad, sickle- or yataghan-shaped distal part with a large lateral incision bordered proximally by a blunt projection, only its outward curved and pointed apical part projecting ventrally beyond margins of sternite 2. Posterior hamuli in lateral view ( Fig. 2 View FIGURE 2 f, 3a) elegant, narrow, tapering to narrowly pointed and slightly curved apices, antero-adaxial margin with a sharp subapical kink; in ventral view ( Fig. 2 View FIGURE 2 h, 3b), posterior hamuli thick and bluntly hooked inside, with a strong subapical incision corresponding to the kink. Vesicle very large, elongate, convex, its anterior part set with fine long hairs. Dorsal side of median segment of penis ( Fig. 4 View FIGURE 4 ) with a large triangular basal and a pair of sickle-shaped lateral membranous parts; median segment of penis with two slanting sclerotised anterior ridges and entirely membranaceous dorsal surface; distal segment of penis with two small, semicircularly curved cornua.
Measurements (mm). Hindwing 23; abdomen without appendages 25; total length (with head and appendages) 36.
Female. Unknown.
Paratype. Similar to holotype ( Fig. 1 View FIGURE 1 b) but differs in the following: Abdomen much shorter and stouter, perhaps because of failure to properly expand. Anterior hamuli ( Fig. 2 View FIGURE 2 g, 3s-d) well seen in lateral view (just a matter of their position) as curving and hooked behind, while the posterior hamuli are somewhat turned along their apices and hence look more strongly curved apically and the subapical kinks of their antero-adaxial margin are hidden in lateral view. Appendages twisted and folded so that cerci diverge slightly, inner arm of one cercus protruding slightly behind main apex (other cercus and epiproct folded). Antenodals 13 (left)–14 (right) on forewings (1st and 5th primary), 9 on hindwings, 1st and 4th (!) primary. Crossveins between Arc and R1-R4 junction 6 above Rs and 5 below Rs on forewing, 4 above Rs and 3 (left)–4 (right) below Rs on hindwing. One cubitoanal crossvein on all wings. Mesepisterna without even traces of antealar spots. Measurements (mm): hindwing 21.5; abdomen 23; total 33.
Short diagnosis. A small (hindwing 21–23 mm) Microgomphus with cerci in dorsal view truncated and slightly bifid, with a blunt angulation of outer margins at 2/3 of their length, strongly curved, sickle-shaped inner arms of cerci arising just behind their middle and not reaching their apices; epiproct arms parallel, closely apposed and only apically slightly diverging; synthorax with broad dorsal pale stripes fused to short mesothoracic collar stripes, isolated from each other, in a 7-like manner and with two parallel dark lateral stripes.
Differential diagnosis. The new species has a unique combination of characters found separately in other species of the genus, but the shape of the inner arms of the cerci is rather specific. The main diagnostic character is the structure of the cerci, with a blunt angulation of outer margins at 2/3 of their length; truncated, slightly bifid apices; and strongly hooked, sickle-shaped inner arms sprouting behind middle of their length and not reaching their apices. The most similar cerci are found in the Sri Lankan Microgomphus wijaya Lieftinck, 1940 , but the inner arms are larger and straight, arising slightly before the middle of the main branch of the cercus; in this species the posterior hamuli are broader ( Lieftinck 1940) and the colour pattern differs strongly (see below). The outer margins of the cerci have slight subapical angulations also in two Indian species, M. torquatus (Selys, 1884) and M. souteri Fraser, 1924 , in which the inner arms diverge before middle and are straight (and very long in the latter species), while the branches of the epiproct are strongly divergent ( Fraser 1934); in other species of the genus the outer margins of the cerci are straight. In M. loogali Fraser, 1923 (Upper Myanmar), the inner arms arise from the middle of the cerci and are curved, although unevenly, but the main branches of the cerci are narrowly pointed and have straight outer margins, while the two apices of the epiproct are strongly divergent ( Fraser 1934). In M. jurzitzai Karube, 2000 (Indochina) , the cerci are similar to those of M. loogali but the inner arms straighter and apices blunter ( Karube 2000). In M. thailandicus Asahina, 1981 ( Thailand) , the cerci are truncated and bifid, the inner arms arise slightly behind the middle of the main branch and are curved but are relatively much shorter than in M. alani , and the main branches of the cerci have their outer margins straight. In addition, in M. thailandicus the epiproct is evenly curved in lateral view and its two apices deeply divided, pointed and nearly parallel, and this species is much larger ( Asahina 1981, 1986). In M. chelifer Selys, 1858 ( Thailand, Cambodia, Malay Peninsula, Borneo, Java, Sumatra), and the few African species of the genus obviously related to it, the long and straight inner arms arise from the cerci subbasally ( Asahina 1990, Orr 2005, Dijkstra 2007, Dijkstra et al. 2014).
Two Asian species of Microgomphus are known only from females, M. (?) verticalis (Selys, 1873) , described from “ India ” (see Fraser 1934) and M. lilliputians Fraser, 1923 , described from “Tennaserim, Burma ” (see Fraser 1934, Kimmins 1966). The new species can be distinguished from them by the synthoracic pattern and size (see below).
As to its coloration ( Fraser 1934, Lieftinck 1940, Asahina 1986, Dijkstra et al. 2014), the new species most resembles M. thailandicus and M. jurzitzai but is much smaller. The dorsal pale stripes on the mesepisternum fuse with the stripes of the mesothoracic collar at their ventral ends to form two inverted ‘7’s, also in M. lilliputians , M. thailandicus and M. jurzitzai ; in other species they either join the collar stripes at their middle ( M. loogali ) or are separated from them ( M. souteri , M. torquatus , M. verticalis , M. wijaya and the African species). The new species lacks or nearly lacks the antealar spot on the mesepisternum, the condition found also in M. lilliputians , M. chelifer and the African species. The new species has two plain black nearly parallel stripes along both lateral sutures of the synthorax (mesepimeron/metepisternum and metepisternum/metepimeron), as in M. thailandicus and M. jurzitzai , while in other species the second stripe is vestigial ( M. loogali ), or the anterior stripe is slanted across the metepisternum and converges with the posterior stripe to form a black ‘Y’ ( M. lilliputians , M. torquatus , M. verticalis and M. wijaya ), or both stripes fuse to each other or nearly so ( M. souteri ), or there is only one (posterior) broad black lateral stripe ( M. chelifer and the African species).
In terms of size (mm) among the Asian species, M. alani (hw 21–23, abd 23–25) is similar to M. chelifer (hw 19–22.5; abd 24), N. torquatus (hw 21, abd 22–24) and M. wijaya (hw 21–23, abd 25–26), larger than M. lilliputians (hw 18, abd 20), smaller than M. souteri (hw 24, abd 29), M. verticalis (hw 25, abd 27) and considerably smaller than M. jurzitzai (hw 26, abd 31), M. loogali (hw 28, abd 31) and M. thailandicus (hw 28–29, abd 33) ( Fraser 1934, Asahina 1981, 1986, 1990, Karube 2000, Orr 2006). However, the existing series of most species are too small to reveal the actual range of variation in size (only M. chelifer is not rare and well collected).
Habitat. The holotype was collected on its maiden flight, at ca 11:30 a.m., in early rainy season, at a small rivulet (‘Macromia rivulet’ in Kosterin 2012, therein reported with somewhat incorrect geographical coordinates) with a muddy bottom, flowing in deep shade in a primary evergreen forest on a hill slope in the Cardamom foothills ( Fig. 5 View FIGURE 5 a). The paratype was collected deeper in the Cardamom foothills in late dry season, also on its maiden flight and near noon, at an extensive broad pool of a medium-sized river ( Fig. 5 View FIGURE 5 b) dividing a primary evergreen forest and a durian plantation and hence a more open habitat. At the type locality, the new species was accompanied by the following, mostly shade-loving forest odonate species: Vestalis gracilis (Rambur, 1842) (several), Euphaea masoni Selys, 1879 (few males), Rhinagrion viridatum Fraser, 1939 (a male), Archibasis viola Lieftinck, 1949 (many males), Pseudagrion pruinosum (Burmeister, 1839) (few males), Idionyx thailandica Hämäläinen, 1985 (a female on her maiden flight), Brachygonia oculata (Brauer, 1878) (several) and, strikingly, Tetrathemis flavescens Kirby, 1889 (two males, a species hitherto known only from Borneo, where is rare; a detailed report of this finding will be published elsewhere); many more species were seen at more open reaches of the same rivulet. The paratype locality was one of the richest for Odonata in Cambodia, sampled during five visits in 2010-2015 in different seasons ( Kosterin 2010, 2011, 2012, 2014, 2015 and unpubl.); other species registered along with the paratype on the same day were Dysphaea gloriosa Fraser, 1938 , Heliocypha perforata (Percheron, 1835) , Pseudagrion rubriceps Selys, 1876 , P. williamsoni Fraser, 1922 , Pseudocopera ciliata (Selys, 1863) , Prodasineura autumnalis (Fraser, 1922) , Neurothemis fluctuan s (Fabricius, 1793), and Trithemis aurora (Burmeister, 1839) ( Kosterin 2010) . The two localities are 30 km apart and are situated at the same moderate elevation of 296 and 341 a.s.l., respectively.
Distribution. The species is so far known from two localities in the southwestern foothills of the Cardamom Mts. in Koh Kong Province of Cambodia. It should also be present in the neighbouring part of the Cardamom SW foothills in Thailand and perhaps in other Cardamomean provinces of Cambodia: Pursat, Kampong Speu, Preah Sihanouk and Kampot. Females reported by Katatani et al. (2010) from Phrae Province of Thailand (see below) could belong to this species as well.




| RMNH |
National Museum of Natural History, Naturalis |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Microgomphus alani
| Kosterin, Oleg E. 2016 |
Microgomphus sp.
| Kosterin et al. 2012: 342 |
| Kosterin 2010: 64 |