
Multitestis paramagnacetabulum, Bray, Rodney A., Cribb, Thomas H. & Justine, Jean-Lou, 2010
|
publication ID |
https://doi.org/ 10.5281/zenodo.294252 |
|
publication LSID |
lsid:zoobank.org:pub:A41D1538-1D85-43CE-9612-50E6A5708ACE |
|
DOI |
https://doi.org/10.5281/zenodo.5629693 |
|
persistent identifier |
https://treatment.plazi.org/id/84E00168-E83A-4E46-84EE-06482D7A3DF7 |
|
taxon LSID |
lsid:zoobank.org:act:84E00168-E83A-4E46-84EE-06482D7A3DF7 |
|
treatment provided by |
Plazi |
|
scientific name |
Multitestis paramagnacetabulum |
| status |
sp. nov. |
Multitestis paramagnacetabulum View in CoL n. sp.
( Figs 9–10 View FIGURES 7 – 10. 7 – 8 )
Type-host: Platax orbicularis (Forsskål) , orbicular batfish (juv) ( Ephippidae ).
Site: Intestine.
Type-locality: Ningaloo Reef, Western Australia (22°42’S. 113°40’E, April, 2000).
Prevalence: 1 of 2 (50%).
Voucher specimens: Holotype QM G 231079, paratypes QM G 231080 – 231085, BMNH 2010.2.10.1-5.
Description: Based on 11 specimens. Measurements in Table 2 View TABLE 2 . Soma almost circular but with slightly protuberant anterior region, short posterior dorsosubterminal ‘tail’. Tegument spinous throughout most of soma; spines minute, not detected on ‘tail’. Oral sucker subterminal, cup-shaped. Ventral sucker large, oval, aperture in form of depressed central region along length of sucker. Prepharynx short. Pharynx subglobular. Oesophagus short. Intestinal bifurcation in posterior forebody. Caeca arcuate, reaching around body parallel to margin, terminating blindly fairly close to posterior extremity of soma, not extending into ‘tail’, termination often obscured by vitelline follicles.
Testes 11, oval, in 2 rosette-like clusters, ventral and overlapping caeca in region between mid-ventral sucker and mid-hindbody, poral field with 6 testes; aporal field with 5. External seminal vesicle saccular, posterodorsal to cirrus-sac. Cirrus-sac claviform, passing transversely dorsal to anterior part of ventral sucker. Internal seminal vesicle oval. Pars prostatica oval or bipartite, lined with anuclear cell-like bodies. Ejaculatory duct long, relatively wide. Genital atrium large, pocketed. Genital pore sinistral, bifurcal, ventral to left caecum.
Ovary multilobate, just posterior to ventral sucker, median. Canalicular seminal receptacle oval, dorsal to ovary. Laurer’s canal not detected. Uterus initially passes posteriorly over ventral surface of ovary, reaches to about mid-hindbody, then passes anteriorly over ventral surface of ovary and into space between ovary and ventral sucker. Eggs relatively few, weakly tanned, operculate. Metraterm muscular, wide, shorter than cirrussac. Vitellarium follicular; dorsal field almost continuous in soma, absent in around edge of worm, anterior to level of pharynx and in median region from mid-ventral sucker posteriorly; not reaching into tail.
Excretory pore dorsally subterminal, on ‘tail’. Excretory vesicle I-shaped, initially narrow, widens in posterior soma, mostly obscured by other internal organs, extent not detected.
Discussion. The worms from Ningaloo are similar to those described by Mamaev (1970) and Bray & Cribb (2003) as Multitestis magnacetabulum , but differ consistently in the testes always being situated a little more posteriorly ( Fig. 9 View FIGURES 7 – 10. 7 – 8 ). Both Mamaev (1970) and Bray and Cribb (2003) stated that the testicular follicles are lateral to the ventral sucker, whereas in the Ningaloo specimens the testes are mostly in the hindbody, although overlapping the ventral sucker. This means that the post-testicular region is relatively shorter in the Ningaloo specimens. Giving dimensions is a problem as the specimens illustrated in detail by Mamaev (1970) lack the distinctive tail-like appendage which always occurs in well-fixed specimens of this species. Mamaev (1970) gives outline illustrations of the shapes taken up by this species, including specimens with ‘tails’, indicating that the fixation method he used often caused contraction or distortion. Nevertheless several dimensions can be quoted from our own measurements (Bray & Cribb 2003 and original data) to indicate the distinctness of the specimens from Ningaloo: pre-testicular distance (34–43% of body length vs 17–27% (our data) and 34–37% (from Mamaev’s illustrations of tailless specimens)), post-testicular distance (22–33% of body-length vs 37–51% (our data) and 21–26% (from Mamaev’s illustrations of tailless specimens)). Measurements of these worms are included in Table 2 View TABLE 2 where they are compared with a series of measurements taken from M. magnacetabulum from Platax teira from Heron Island and New Caledonia. The specimens from Ningaloo, Heron Island and New Caledonia were fixed in the same way, so the slight differences found can be considered genuine local variation.
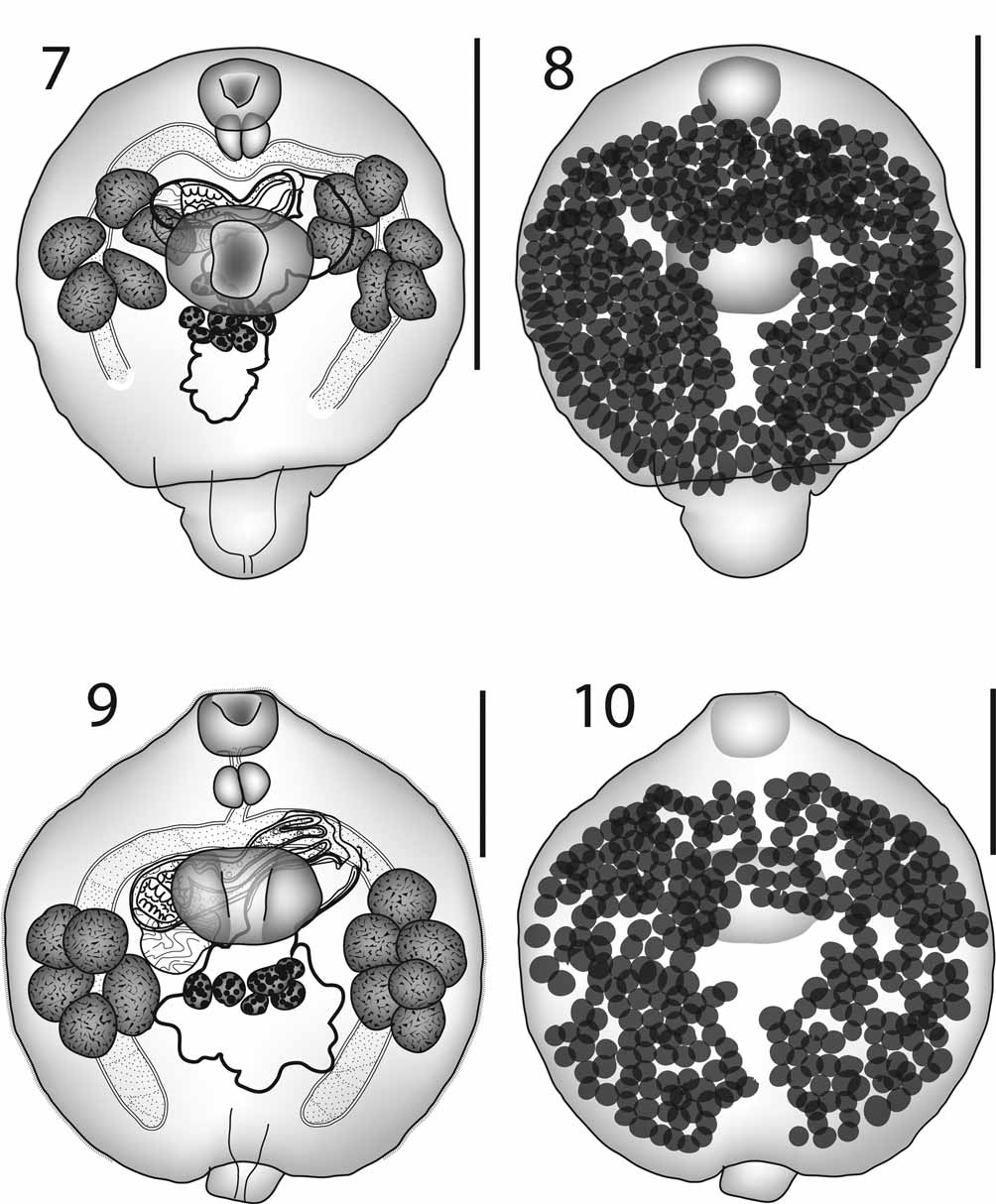
TABLE 2. Dimensions and ratios of Multitestis magnacetabulum and M. paramagnacetabulum n. sp.
| Species | Multitestis magnacetabulum | Multitestis magnacetabulum | Multitestis paramagnacetabulum n.sp. |
|---|---|---|---|
| Host | Platax teira | Platax teira | Platax orbicularis |
| Locality | Heron Island | New Caledonia | Ningaloo |
| n | 18 | 1 | 11 |
| Length | 636–1,351 (874) | 781 | 520–625 (603) |
| Width | 570–957 (722) | 748 | 419–621 (500) |
| Forebody | 217–366 (277) | 295 | 176–219 (202) |
| Pre-oral lobe | 8–18 (11) | 7 | 0–5 (2) |
| Oral sucker | 98–148 × 118–168 (114 × 140) | 136 × 155 | 68–92 × 85–109 (75 × 97) |
| Prepharynx | 0 | 0 | 7–21 (15) |
| Pharynx | 57–99 × 60–103 (74 × 80) | 100 × 122 | 45–60 × 55–71 (52 × 63) |
| Oesophagus | 0–17 (2) | 0 | 0–28 (11) |
| Intestinal bifurcation to ventral sucker | 64–172 (118) | 90 | 0–56 (15) |
| Vitellarium to ventral sucker (V-VS) | 117–221 (163) | 201 | 74–132 (92) |
| Ventral sucker (VS) | 141–225 × 192–318 (175 × 250) | 182 × 267 | 87–142 × 141–186 (103 × 157) |
| Cirrus-sac | 205–347 × 58–140 (255 × 86) | 274 × 82 | 196–293 × 60–88 (253 × 75) |
| Extent of external seminal vesicle into hindbody (EESSH) | 0 | 0 | 0–78 (35) |
| VS to Ovary | 0 | 0 | 0–36 (9) |
| Ovary | 88–184 × 102–219 (126 × 162) | 98 × 132 | 46–131 × 83–149 (88 × 120) |
| Testicular field Distance between testicular fields | 222–407 × 164–330 (290 × 228) 102–275 (155) | 254–276× × 226–236 158 | 151–229 × 99–205 (186 × 147) 115–232 (147) |
| Post-testicular region | 247–642 (383) | 301 | 121–207 (158) |
| Post-caecal length | 120–310 (215) | ? | 146–159 (152) |
| Eggs | 49–58 × 20–31 (54 × 25) | 52 × 27 | 49–62 × 28–39 (57 × 34) |
| Width as % of body-length | 71–94 (84) | 96 | 78–93 (83) |
| Forebody as % of body-length | 27–37 (32) | 38 | 30–39 (34) |
| Sucker-length ratio | 1:1.36–1.70 (1.53) | 1:1.3 | 1:1.2–1.9 (1.4) |
| Sucker-width ratio | 1:1.6–1.9 (1.8) | 1:1.7 | 1:1.5–1.8 (1.6) |
| Pharynx: oral sucker ratio | 1:1.5–2.0 (1.8) | 1:1.3 | 1:1.4–1.6 (1.5) |
| VS-Ovary as % of body-length | 0 | 0 | 0–5.4 (1.5) |
| EESVH as % of VS-Ovary | N/A | N/A | N/A |
| Post-testicular region | 37–51 (43) | 39 | 22-33 (26) |
| Cirrus-sac length% of body-length | 25–34 (30) | 35 | 36–46 (42) |
| Pre-vitelline distance % of body- length | 10–17 (13) | 12 | 14–22 (18) |
| Testicular field length% of body- length | 30–38 (34) | 33–35 | 24–35 (31) |
| Pretesticular distance% of body-length | 17–25 (22) | 27 | 34–43 (38) |
| Post-uterine distance% of body-length | 22–40 (30) | 20 | 17–29 (24) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.