
Hyphinomos svenhedini Ramme, 1950
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5099.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:E06557A6-CDD9-4077-9C76-69C14960476C |
|
DOI |
https://doi.org/10.5281/zenodo.6310509 |
|
persistent identifier |
https://treatment.plazi.org/id/03DD87B8-FFCD-4425-FF04-F983FF0FE5F5 |
|
treatment provided by |
Plazi |
|
scientific name |
Hyphinomos svenhedini Ramme, 1950 |
| status |
|
Hyphinomos svenhedini Ramme, 1950 View in CoL
Figures 1 View FIGURE 1 , 2B View FIGURE 2 , 3A–C View FIGURE 3 , 4–14 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 View FIGURE 12 View FIGURE 13 View FIGURE 14
Hyphinomos svenhedini Ramme, 1950: 76 View in CoL ; Jin and Xia, 1994: 30; Sergeev, 1995: 218; Braun, 2016: 104.
Type locality: Indian subcontinent, Trans Himalaya, Ladakh, Indus valley north of Hanle.
Type material (all NMW): Holotype — 1♂, Allotype — 1 ♀, both Hanle, Ladakh. Paratype — 1 ♀, Lahaul [Lahoul], Himachal Pradesh .
Material studied: Type series deposited in NMW. 3 ♂ and 2 ♀, India: Ladakh , Kargil , Shahsi lake, 80km from Kargil, ~ 4206 m a.s.l., 34°33’57.12”N 76°21’25.74”E, 13.viii.2020 and 25.viii.2021 collected by A. Ali and identified by R. Jaiswara & K. G. Heller GoogleMaps .
Translation of Ramme’s (1950) description (references to the figures of the present paper)
Head. Head rather short and thick; face vertical ( Fig. 6E View FIGURE 6 ), slightly wrinkled; cheeks very broad and pronounced ( Fig. 6E View FIGURE 6 ); male with arched occiput ( Fig. 5D View FIGURE 5 ), female flatter, smooth ( Fig. 5B View FIGURE 5 ), 2 longitudinal rows of fine points on both sides of the midline. Antennae somewhat longer than the abdomen ( Fig. 5B View FIGURE 5 ); dense and finely haired, especially in the apical two-thirds, the basal segment strongly thickened ( Fig. 2B View FIGURE 2 ). Eyes small, spherical broadly oval ( Fig. 5 View FIGURE 5 & 6A, D, E View FIGURE 6 ); lower margin in the same line with antennae-bases ( Fig. 6E View FIGURE 6 ). Fastigium vertex convex ( Fig. 5B, D View FIGURE 5 ), with slightly angular side margins, rounded in front, with flat, but sharply defined longitudinal furrow in the basal half or up to two thirds, profile convex.
Pronotum. Back of pronotum roughly square ( Fig. 6D View FIGURE 6 ), prozona with bulging lateral lobes laterally ( Fig. 6A & D View FIGURE 6 ); steeply sloping in metazona ( Fig. 5B & D View FIGURE 5 ), anterior transverse furrow consistently sharp, concave ( Fig. 5A, B View FIGURE 5 & 6A, D View FIGURE 6 ); V-shaped middle transverse furrow interrupted in the middle, posterior transverse furrow parallel to pronotum posteriorly ( Fig. 6A & D View FIGURE 6 ). Metazona somewhat rising, more finely wrinkled than prozona ( Fig. 5B & D View FIGURE 5 ); anterior margin slightly concave ( Fig. 5A, C View FIGURE 5 & 6A, D View FIGURE 6 ); central keels are missing. Lateral lobes very uneven, wrinkled, broadly deepened along the lower margin, set off in bulges against the metazoan, front edge of the lobes short, broadly rounded at obtuse angles, merging into the somewhat concave rear edge ( Fig. 5B & D View FIGURE 5 ). Prosternum with 2 small, widely spaced tubercles, mesosternum with rounded lobes at right angles, metasternum with broad, short, apically somewhat thickened and convexly rounded ridges.
Wings. Male with short tegmina not reaching end of 1st tergite, broadly rounded apically ( Fig. 5C View FIGURE 5 & 6B View FIGURE 6 ); female laterally, rounded ( Fig. 5A View FIGURE 5 & 6A View FIGURE 6 ), in both with thin veining.
Legs. Fore coxae with strong, flat, slightly downwardly curved spines. Fore femora fairly broad, without spines, fore tibiae somewhat constricted behind the rather thickened part of the auditory organ, with a terminal spine on the top outside, with 6 spines each on the outside and inside edge on the underside, as are the central splints. Hind femora as long as, or not quite as long as, twice the length of the middle femora, rather thick in the basal half, lower inner edge with 4–8 spines, outer edge with 5–10 spines (opposite [H.] fasciata Uv. not only in the apical half); Vertical rear carina broad, in the apical half sometimes slightly curved downwards, all keels obtuse, on the inside with 8–13 on the upper side, 10–14 on the outside, on the inside with 4–5 on the underside, on the outside 5–6 spines; lower pair of terminal spines considerably more delicate than the upper, hardly half as long as this one. All tarsi depressed, on the sides rather strongly furrowed, plantulae very short, shorter than the terminal spine, quite thick and blunt, immobile.
Abdomen. Male anal tergites narrow, transverse; longitudinal pit in the apical half ( Fig. 5C View FIGURE 5 ), 2 flat tubercles at the base, slightly incised at middle at the rear edge ( Fig. 8C View FIGURE 8 ). Anal plate (not visible in situ because it is strongly pointing downwards) briefly tongue-tied. Cerci hidden under the anal tergite, short, with claw-shaped (4–5 teeth), curved inward inner part and short blunt outer end part ( Fig. 9 View FIGURE 9 & 10A View FIGURE 10 ). Subgenital plate transverse, margins broad and rounded, slightly incised in the middle; styli short, thick and blunt, directed inwards ( Fig. 8D View FIGURE 8 ). Female anal tergite at the rear edge on both sides of the middle with a slight adherence; anal plate short, tongue-shaped, with a lighter central furrow, thick at the base, apically pointed. subgenital plate transverse, with broad lateral marginal ridges, slightly incised in the middle ( Fig. 8E View FIGURE 8 ). Ovipositor saber-shaped, slightly curved upwards, thick at the base, at apical point; upper and lower margin finely wrinkled ( Fig. 3A–C View FIGURE 3 ).
Color. Glossy black; antennae blackish, light brownish at base ( Fig. 5 View FIGURE 5 ). The face between the clypeus and the antennae bases is ivory in color along the narrow blackish clypeus ( Fig. 4B View FIGURE 4 & 6E View FIGURE 6 ) (thus reminiscent of Drymadusa and relatives). Occiput and pronotum black ( Fig. 4 View FIGURE 4 , 5C View FIGURE 5 & 6B View FIGURE 6 ), elytra ivory colored ( Fig. 4 View FIGURE 4 , 5 View FIGURE 5 & 6A & B View FIGURE 6 ). Back and sides of the abdomen brown-black, the last 4 tergite above ivory color ( Fig. 4 View FIGURE 4 & 5C–D View FIGURE 5 ), belly whitish brownish. Front and middle thigh, rails dirty yellowish gray ( Fig. 4 View FIGURE 4 & 5C–D View FIGURE 5 ). Hind leg outside in the basal third of the same color, otherwise black ( Fig. 4 View FIGURE 4 ), remainder and tarsi pure light brown, only slightly narrow near the knee blackened ( Fig. 4 View FIGURE 4 ). Face, occiput, pronotum and abdomen ridge olive-brown to dirty rust-brown. Brown black side lobes of pronotum ( Fig. 5B & D View FIGURE 5 ). Ovipositor monochrome light brown ( Fig. 3A–C View FIGURE 3 ).
Emended description. Entire body covered with fine distantly spaced setae. Head. Face in front view almost 1.5 times longer than wide; frons bearing a pair of shallow vertical furrows ( Fig. 6E View FIGURE 6 ). Fastigium as wide as scape. Scape as long as wide. Epistomal suture angular; subgenal suture arched. Ocelli small, disposed in triangle, lateral ocelli oval, more dorsal, present on the lateral margin of protruded fastigium, slightly above the upper margin of antennal socket; median ocellus round, almost in the line of basal margin of eyes and antennal socket; distance between median and lateral ocelli twice of distance between lateral ocelli. Maxillary palpi 5 th article longest, 3 rd smaller than 4 th article; 5 th article slightly widening and truncated distally ( Fig. 6C & E View FIGURE 6 ). Legs. FI ventral side 1–2 spines on outer margin, no spines on inner margin; no spine on dorsal side. TI with tympanum, in dorsal view appears as cup-shaped, bulging out from the surface of tibia ( Fig. 6F View FIGURE 6 ); dorsal side inner margin with 1 spur, nothing on outer margin; ventral side of inner and outer margin with 6 spurs. FII ventral side 1–2 spines on both inner and outer margin (sometimes no spines on inner); no spine on dorsal side. TII dorsal side inner margin with 1 spur, unarmed on outer margin; ventral side of inner and outer margin with 6 spurs. Proximal part of FI and FII almost 1.5 time wider than distal. FIII proximally almost 3 times wider than distal; FIII ventral side 4–6 spines on inner margin, 4–9 spines on outer margin; no spine on dorsal side. In males (n=3), TIII ventral side inner margin 4–6 spurs, outer 6–7 spurs; dorsal side inner margin 6–8 and outer 3 spines. In females (n=2), TIII ventral side inner margin 4–5 and outer 6–7 spurs; dorsal side inner margin 12–14 and outer 10–12 spines.
Male. Tegmina overlapping; dorsal field with distinct anal veins, anterior cubital and median vein, no distinct harp and mirror; apical and lateral field completely reticulated ( Fig. 6B View FIGURE 6 ). Ventral side of elytra: stridulatory file on PCuA with approximately 121 teeth ( Fig. 7A View FIGURE 7 ). Shape of tooth elongated, medially ridged, fine transverse furrows along the length ( Fig. 7B View FIGURE 7 ), average inter-tooth distance in the middle of the file is 10.6 µm (±0.8 µm; n=14).
Male genitalia. Cerci with inner part claw-shaped bearing 3–5 teeth. These teeth are quite similar in both left and right side cerci of an individual, but surprisingly variable between different males ( Fig. 9 View FIGURE 9 & 10A View FIGURE 10 ). Titillators with one pair of very massive sclerites, strongly curved, basal branch longer than distal branch which carries small spines on both edges ( Fig. 10B & C View FIGURE 10 ).
Female. Pronotum and abdomen olive green in color; the last 4 tergite somewhat ivory ( Fig. 4C View FIGURE 4 ). Tegmina non-overlapping and distantly spaced, anterior cubital and median vein distinct, dorsal and lateral field completely reticulated ( Fig. 5A View FIGURE 5 & 6A View FIGURE 6 ).
Female genitalia. [in Ramme] Supra anal plate short, V-shaped, styli long ( Fig. 8F View FIGURE 8 ).
Color. Head. Scapus black, 2 nd –7 th antennal segment light brown. In female, dorsal side of head behind fastigium, either yellow interspersed with black spots (in freshly molted adult) or black and adorned with yellow stripes and patches ( Fig. 2B View FIGURE 2 & 4C View FIGURE 4 ). Mouth parts ivory, maxillary palpi- distal margin of 5 th, proximal margin of 2 nd and 3 rd brown and 1 st article in male brown, in female only proximal margin brown. Mandible light brown with peripheral margin blackish-brown ( Fig. 6C View FIGURE 6 ). Pronotum. In male, dorsal lobe of pronotum same as lateral lobe ( Fig. 6B View FIGURE 6 ); in female, dorsal lobe light brown similar to occiput, sometimes interspersed with black patches ( Fig. 6A & D View FIGURE 6 ). Tegmina ivory in both male and female; in male proximally brownish. Legs. Foreleg and midleg mostly pale yellow; in males FI ventrally brown towards distal end. FIII exterior surface with short vertical brown lines; both in male and female half of FIII brown to blackish brown. TI-TIII pinkish ( Fig. 4A–C View FIGURE 4 ). Male subgenital plate ivory with lateral margins blackish-brown; supra anal plate proximal side blackish-brown ( Fig. 8C & D View FIGURE 8 ).
Note: Coloration best seen in individuals when present in natural habitat. Wet preserved specimens slightly discoloured with time.
Measurements. Morphometric measurements of H. svenhedini Ramme, 1950 type series and specimens used in this study are listed under Table 1 View TABLE 1 .
Mating behaviour. Like many other insects in alpine zones of high mountains ( Mani, 1968) and high altitude crickets ( Gurney & Liebermann, 1975), this species is also diurnally active hence mating takes place in the day time. September month seems to be the peak season for them, hence mating pairs were seen everywhere in the areas around Shashi lake. It was an unmistakably common sight.
Mating behaviour of this species was studied mostly in the laboratory. A nylon mesh cage of 12”x12”x12”(30 cm) dimension was used to enclose a male and a female together. Soon after releasing them together, both the sexes were found to be toppling over each other alternately wherein male was trying to attain the copulatory position by frequently climbing over female’s back. In doing so, male was found to be rubbing his forewings briefly for a couple of seconds to produce high frequency courtship signal. Unlike other orthopterans, attraction and coupling via antennation was absent and pair formation was not preceded by acoustic signalling in this species. Males either leaps abruptly onto the female’s body or simply climbs over her back and holds her abdomen with the help of his legs to restrict her movement since female was always in the process of resisting male and moving away. Males and females exhibit an end-to-end copulation position ( Fig. 12 A&B View FIGURE 12 ) as depicted by Alexander & Otte, 1967 ( Fig. 11–12 View FIGURE 11 View FIGURE 12 ). The final copulatory position was achieved by the male by rapidly repositioning himself by bending abdomen in a c-shape underneath the female, his head facing the ovipositor and then using his pincer like cerci to grip the soft cuticle of the underside of the female’s abdomen and restricting her movement with his mid legs. He would grip the ovipositor with the help of foreleg only for first few minutes (2–4 minutes), eventually the grip of cerci becomes so strong that it allows the male to leave the grip of his foreleg from ovipositor. Apparent resistance by female to copulation was observed where female was seen walking rapidly during mounting and kicked male with her hind legs. On the contrary, females in some cases tend to start copulation and males resist by jumping.
Copulation lasted for approximately one hour fifteen minutes in one couple and on separation no spermatophore was found attached to the females abdomen. In an attempt to observe the spermatophore size in this species, copulation between another mating couple was paused after one hour however, no spermatophore was seen. In a third mating pair, copulation lasted for approximately 3.5 hours, suggesting duration of copulation in laboratory condition to be variable. We do not have data on duration of copulation in their natural habitat. However, concluding from the high frequency of mating pairs observed in the field, copulations may either last long or happen quite often or both.
Acoustic signals: The calling song is composed of a group of short and long series of impulses where the long series (assumed to be closing hemisyllables) are preceded by shorter ones (assumed to be opening hemisyllables; Fig. 13A–B View FIGURE 13 ). Number of syllables vary between 5–6 per echeme. Duration of short and long hemisyllables are 13 ±1 ms and 41 ±8 ms (mean ± SD), respectively. Period from the onset of short to the onset of consecutive hemisyllable is 23 ± 9ms, from the onset of long to the onset next hemisyllable 56 ± 5ms. Duration and periods of syllables are 65 ±13 ms and 81 ±10 ms resulting in a syllable repetition rate of 12 Hz (at approximately 17.5 ºC; average indoor temperature on 16/09/ 2021 in Kargil). Duration and period of echemes are 406 ±36 ms and 2692 ±789 ms, respectively. The dominant recorded frequency of calling song is 16.8kHz, but may be limited by the recording device ( Fig. 13C View FIGURE 13 ; compare Fig.14C View FIGURE 14 ).
Signal produced during mounting attempts by male is composed of separated syllables, differing in structure from that of the calling song ( Fig. 14A–B View FIGURE 14 ) and peak frequency lies between 30–35kHz ( Fig. 14C View FIGURE 14 ).
Distribution and habitat: This species is one of the high-altitude Ensiferans found in India. Ramme (1950) documented one male paratype and a female allotype collected by the famous biologist Ferdinand Stoliczka (see https://en.wikipedia.org/wiki/Ferdinand_Stoliczka) during his first Himalaya expedition in August 1864. The type specimens were collected from an altitudinal range of 4572–5486 m (a.s.l) from Indus Valley, North of Hanle situated in the valley of Hanle river which originates from Imis La pass on the border of India-Tibet and joins the Indus River at Loma, close to the town of Nyoma. Another female collected from Lahaul (approx. 4267 m altitude a.s.l, as read from map) (in the collection of Brunner von Wattenwyl: “ex Mus. Lübeck”) was designated as paratype by Ramme (1950) ( Fig. 1 View FIGURE 1 ).
Until 2016, the species was known only from its type specimens as described by Ramme 1950. Another specimen from Zanskar, 3800 m a.s.l., 26 viii 2016, 33° 59’ 59.37’’ N 76° 46’ 15.98’’ E came into documentation based on a male photograph taken by Christine Provot ( Fig. 1 View FIGURE 1 ) on the website- https://www.galerie-insecte.org/galerie/ref-174389.htm.
The specimens studied here were collected from area around the Shashi Lake ( Fig. 1 View FIGURE 1 & 11A–B View FIGURE 11 ), located at an altitude of 4206 m a.s.l., 80 km away from Kargil district towards north east. The lake was built by the villagers of Sandow approximately three decades ago for the storage of water and named ‘Shashi Tso’ (Shashi=Mountain, Tso=lake) and ‘Purgi’ in their local language. In winter, the lake gets frozen and remains covered by snow. Towards the end of March, snow from adjoining hills melts and the water flows down into the lake.
During the months of April to October, the mountain and surrounding areas of the lake thrive with a variety of economically significant shrubby plants ( Fig. 11C–E View FIGURE 11 ) ( Table 3 View TABLE 3 ). As the temperature during these months ranges between 14.1°C to 26.9°C ( Tundup et al., 2016) several noteworthy animals ( Table 2 View TABLE 2 ) also become active and are sighted.






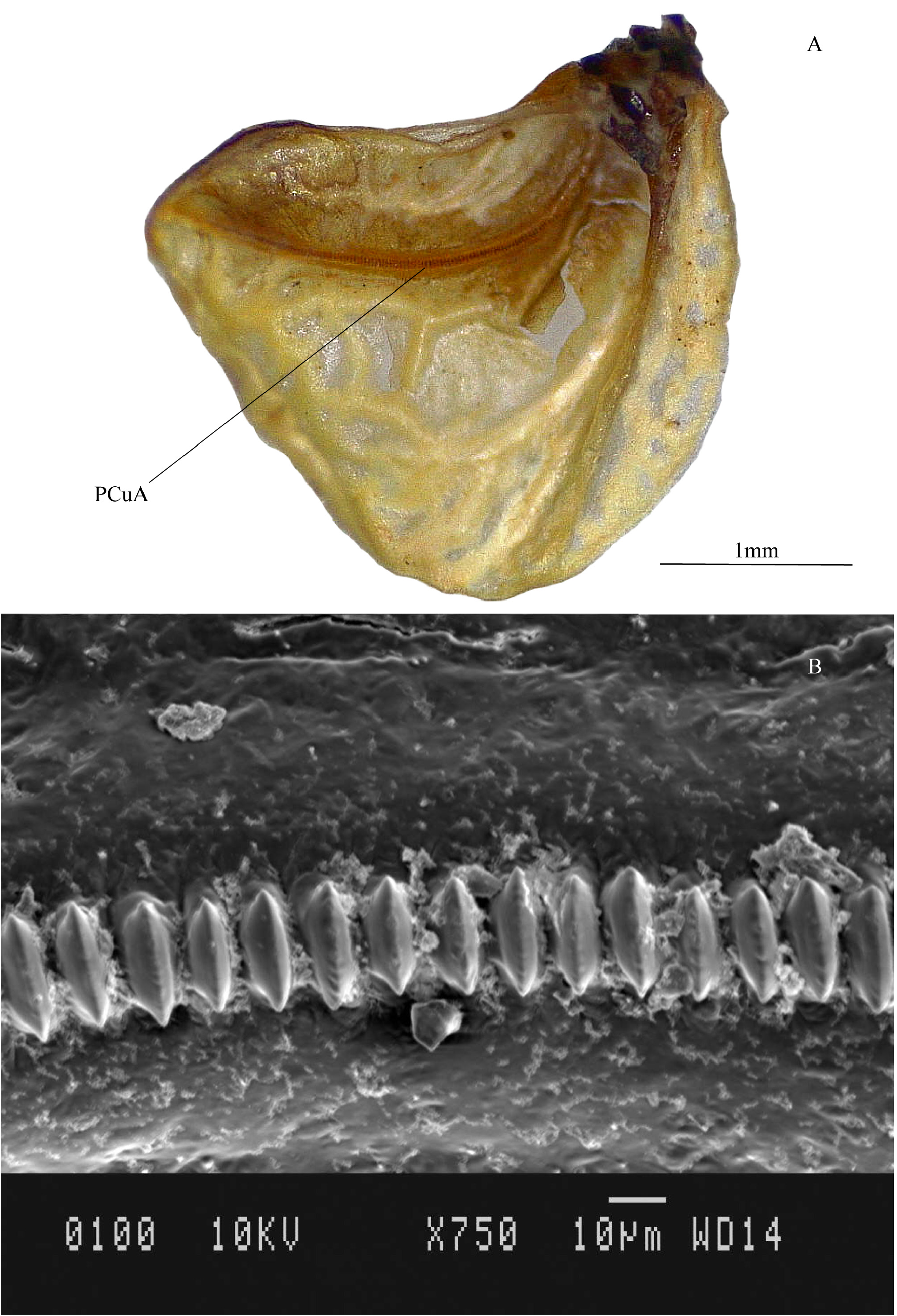


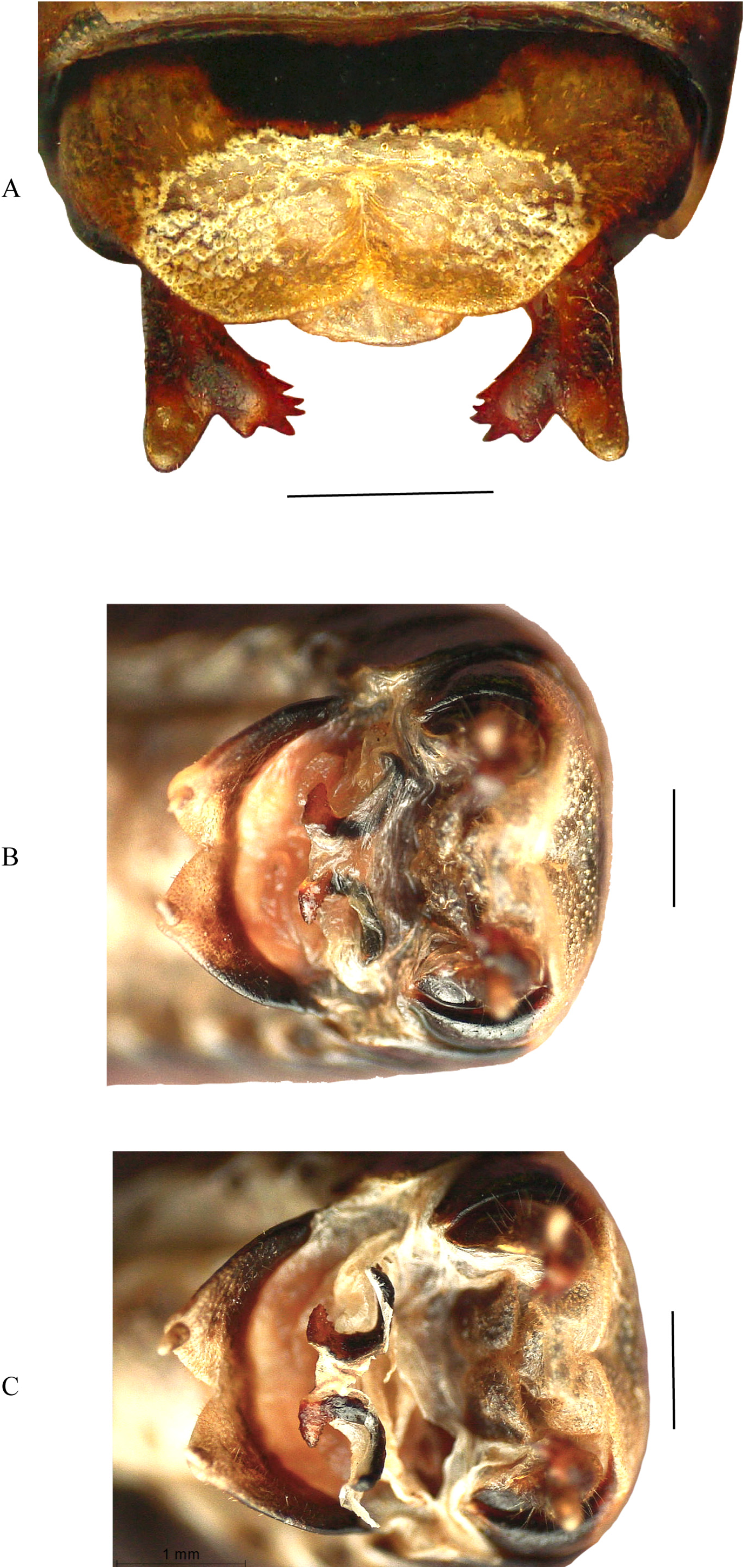




TABLE 1. Measurements of H. svenhedini type series and specimens used in this study. Abbreviations as listed in material and method section.
| Males | BL | PL | PW | FW_L | FI_L | FIII_L | TIII_L | |
|---|---|---|---|---|---|---|---|---|
| Holotype | 14–15 | 3.5–3.9 | – | 2.5 | 4.4–4.7 | 10.5–10.7 | 10.5 | |
| M1_Wet | 18.2 | 3.4 | 4.7 | 2.4 | 4.1 | 10.1 | 9.7 | |
| M2_Dry | 14.8 | 3.3 | 4.5 | 2.5 | 4 | 9.9 | 9.7 | |
| M3_Dry | 14.9 | 3.6 | 4.5 | 2.5 | 4 | 10.3 | 9.6 | |
| Females | BL | PL | PW | FW_L FI_L | FIII_L | TIII_L | Ovi_L | |
| Paratypes | 17.5–18 | 4.5–4.7 | – | 1.5 | 5 | 12–12.5 | 11.5 | 11.7–12 |
| F1_Wet | 19.1 | 4.1 | 5.3 | 1.9 | 4.4 | 11.2 | 10.4 | 12.7 |
| F2_Dry | 15.3 | 4 | 5.2 | 1.6 | 4.5 | 11.8 | 10.4 | 12.7 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Tettigoniinae |
|
Genus |
Hyphinomos svenhedini Ramme, 1950
| Ali, Ahsan, Jaiswara, Ranjana, Bharti, Umesh & Heller, Klaus-Gerhard 2022 |
Hyphinomos svenhedini
| Braun, H. 2016: 104 |
| Sergeev, M. G. 1995: 218 |
| Jin, X. - B. & Xia, K. - L. 1994: 30 |
| Ramme, W. 1950: 76 |