
Telosticta, Dow & Orr, 2012
|
publication ID |
https://doi.org/10.5281/zenodo.5348617 |
|
persistent identifier |
https://treatment.plazi.org/id/03DD87E5-FFEF-6157-FECB-F917FDFE99D5 |
|
treatment provided by |
Tatiana |
|
scientific name |
Telosticta |
| status |
gen. nov. |
Telosticta View in CoL , new genus
Type species. — Protosticta feronia Lieftinck, 1933 View in CoL by present designation
Diagnosis. — A genus of slimly built platystictids ranging in body length from medium to long. Ground colour generally dark, always with pale antehumeral markings and some pale markings on at least one of abdominal segments 8–10. Males are readily distinguished from all other members of the Platystictidae by the combination of a narrow vertex, ventrally projecting, flattened lateral processes on the posterior pronotal lobe, pale antehumeral markings and terminal segment of penis with a well-developed convex tongue-like structure between the horns. The superior anal appendages are typically down-turned in apical one half to one third, with a process or bulbous swelling arising dorsally and interiorly from the basal one-quarter to one half, and another, often poorly developed, arising dorsally at the down-turn. Inferior anal appendage with an internal subapical spine and its apical part modified into a dorsal-ventrally flattened structure that is typically upwardly concave and scoop-like in appearance.
Etymology. — Telosticta a feminine noun, compounded from the Greek τ λος, meaning toll, named for Jan van Tol, whose surname originates from the Dutch word of the same meaning, in honour of his contributions to our knowledge of the Platystictidae , and the suffix – sticta, a common generic ending in the family.
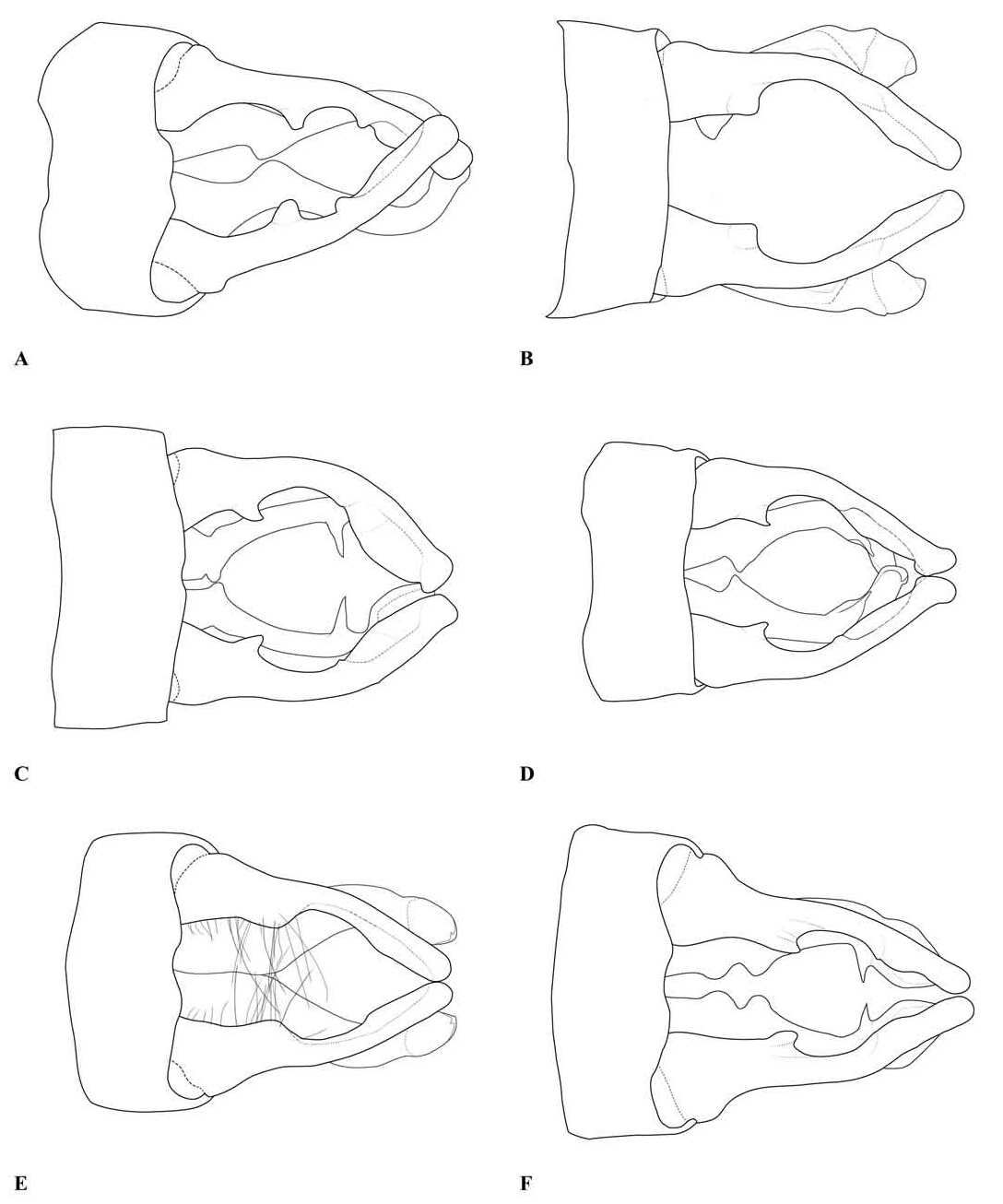
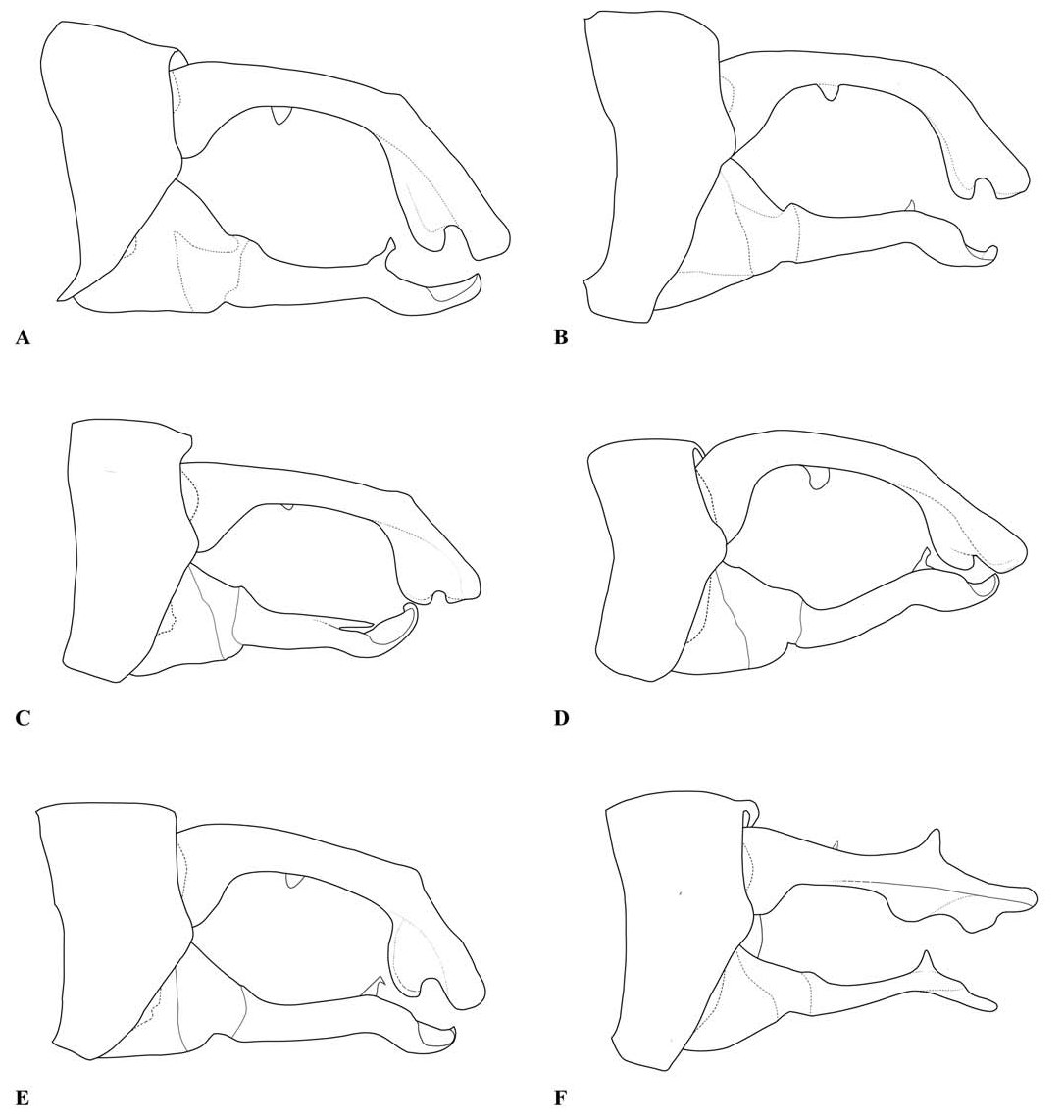
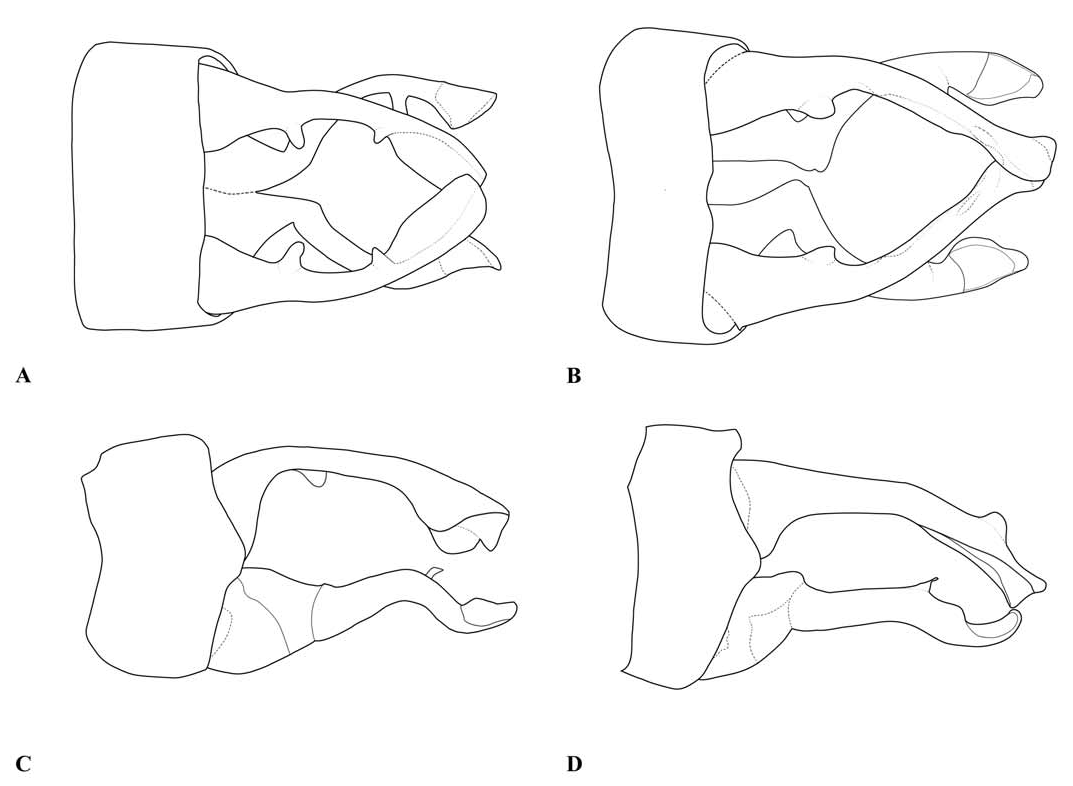
Description. — Head (e.g., Fig. 1A View Fig ): mostly black, with much of the labrum, the anteclypeus and part of the mandible bases pale. The dorsum of the head is narrow, with the ratio of the width of a compound eye measured at the narrowest part of the vertex to the distance between compound eyes at the same point typically around 9/10 (ca. 0.85–0.96 in specimens measured, including at least one example of each species included in the genus). The transverse anterior carina is typically well developed, with its lateral extremities angulated and often prominent.
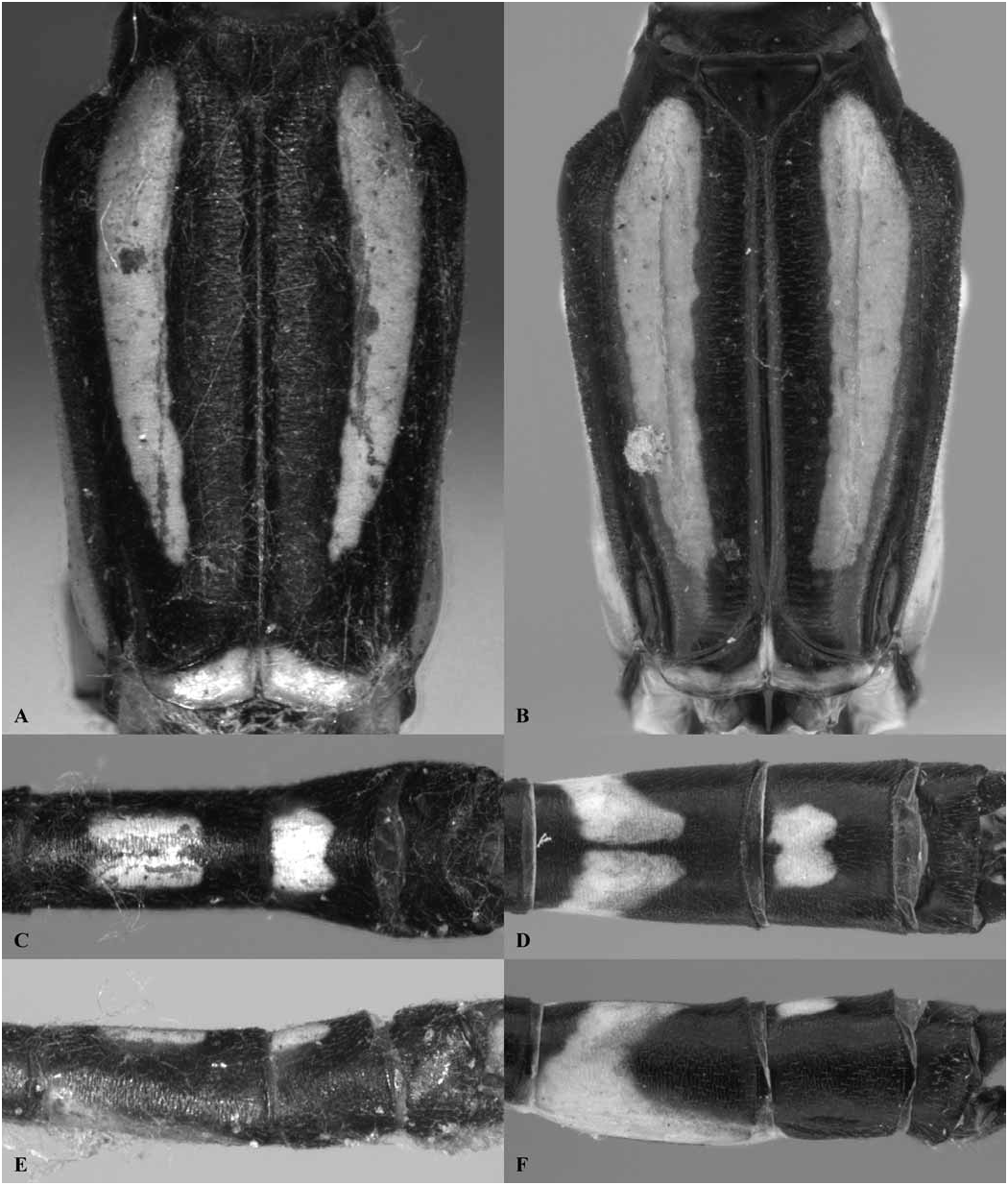
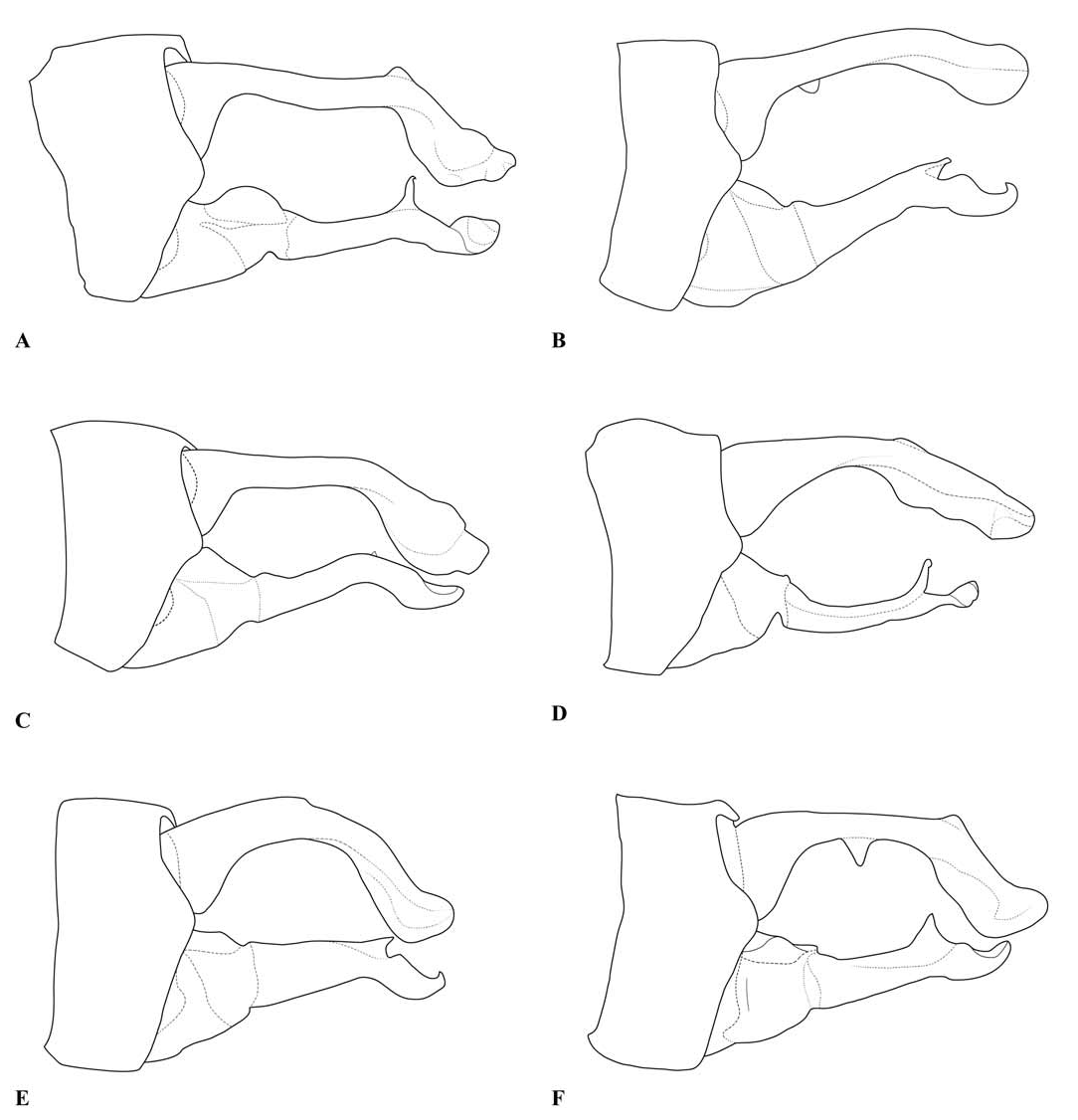
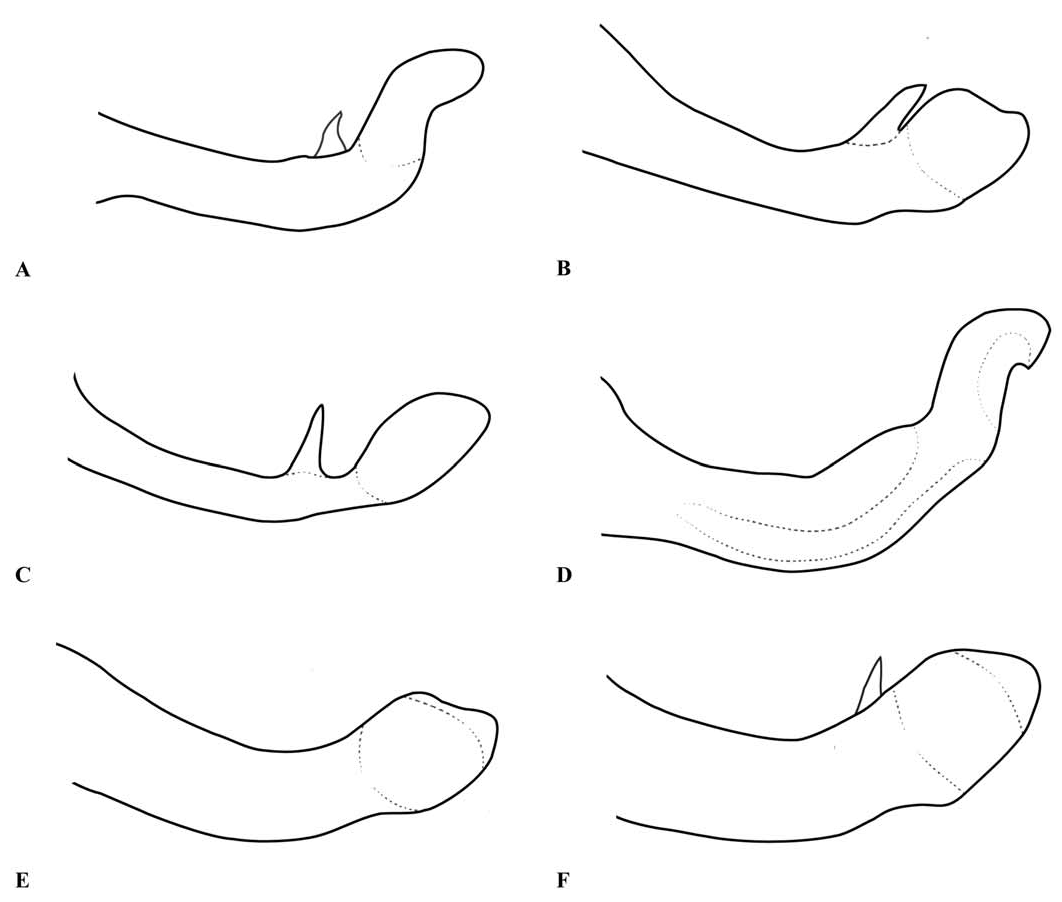
Thorax: prothorax typically mostly pale, darker to the rear of the propleuron and the middle pronotal lobe. The posterior pronotal lobe is black; its hind margin is normally simple in both sexes. In the male the posterior lobe bears at each side a flattened lateral process, the “remarkable pending appendages of the posterior lobe of the pronotum” of van Tol (2005), directed downwards beside the rear margin of the propleuron and typically expanding slightly from its origin to the tip, which is normally rounded (e.g., Figs. 3 View Fig , 9 View Fig , 17A, B View Fig ). The lateral process is variably developed in females even of the same species, and sometimes absent in that sex. The synthorax (e.g., Figs. 1B View Fig , 4 View Fig , 10 View Fig , 16A, B View Fig ) bears pale antehumeral markings on the mesepisternum, which is otherwise bronzy black. The mesepimeron is typically black and the metepisternum is dark with a large pale marking. The metepimeron is largely or entirely pale and the venter of the synthorax is pale. The legs are mostly pale. The wings are falcate, with the arculus distal to Ax2 and its sectors not separated until some distance from origin. The quadrilateral is long and narrow and nearly rectangular. The anal bridge (ab) is variably present, in some species it is absent (e.g., traditional Protosticta venation), in others present (e.g., traditional Drepanosticta venation), and while each species typically has ab either present or absent, it is not uncommon to find individuals which have the other form in from one to all four wings. In one species ( T. belalongensis ) even the anal crossing (ac) is absent or vestigial in three wings of the single specimen available. When ab is present ac typically arises at ca the level of Ax2, when ab is absent ac typically arises proximal to Ax2, but closer to Ax2 than Ax1. R 4 arises at or distal to the subnodus, IR 3 well distal to the subnodus. The pterostigma typically covers slightly more than one underlying cell, and is dark brown or black in mature individuals, often with a pale border.
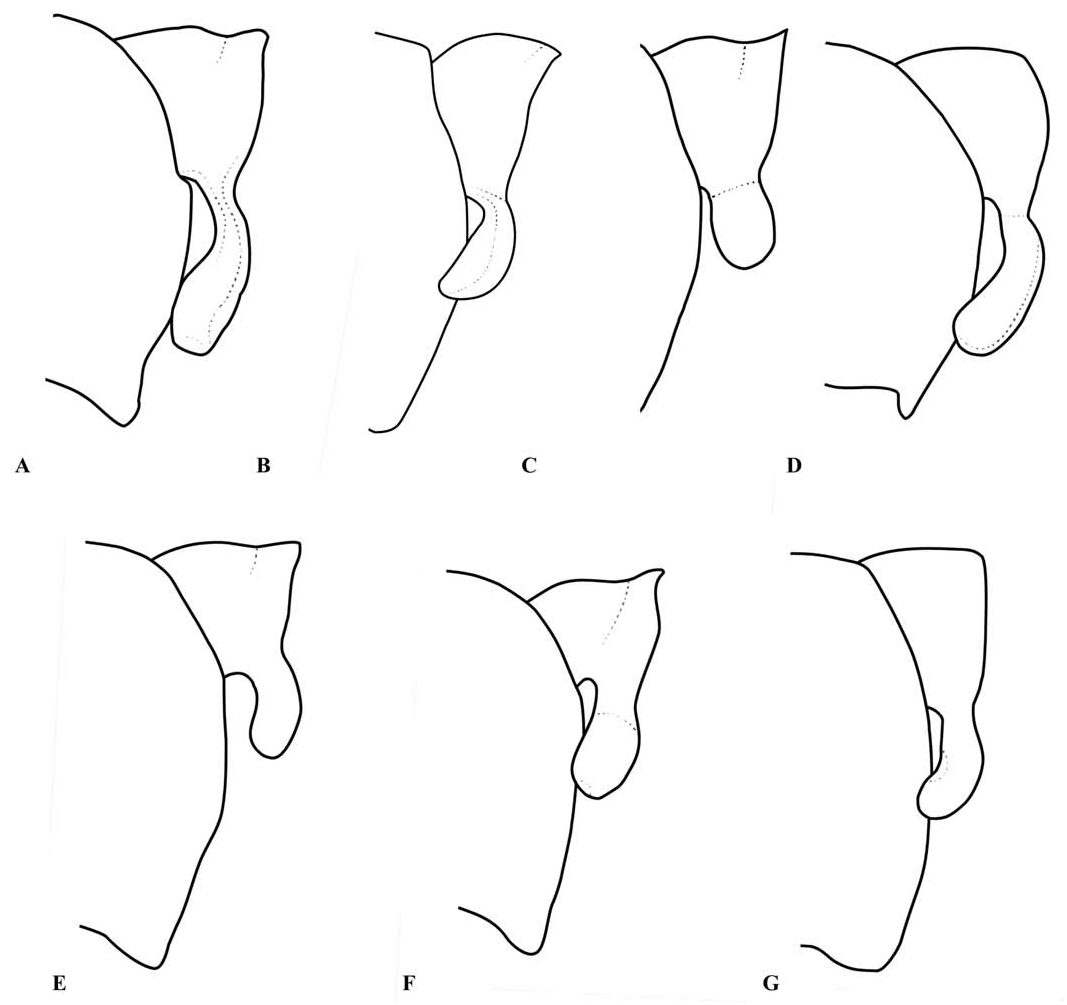
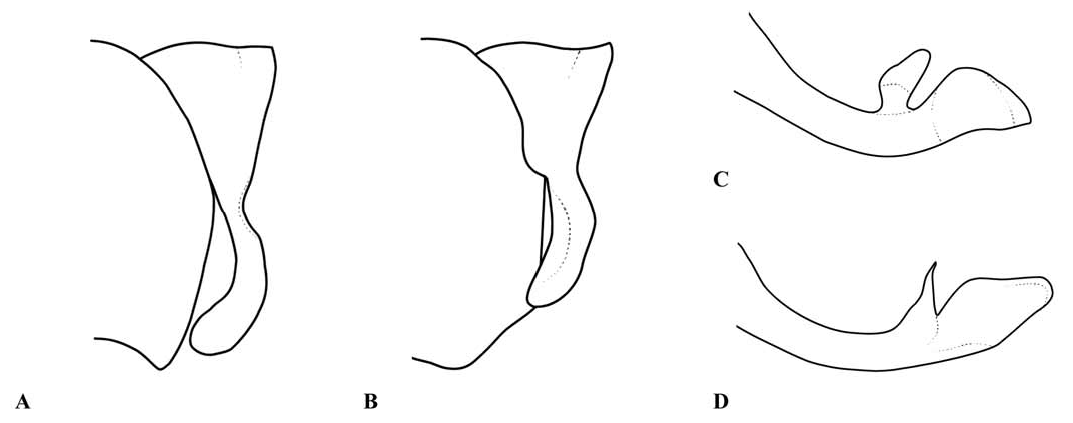
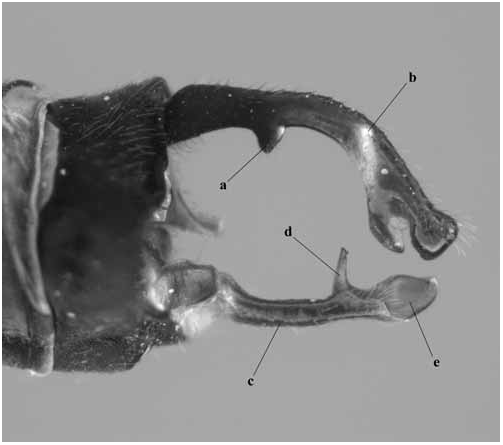
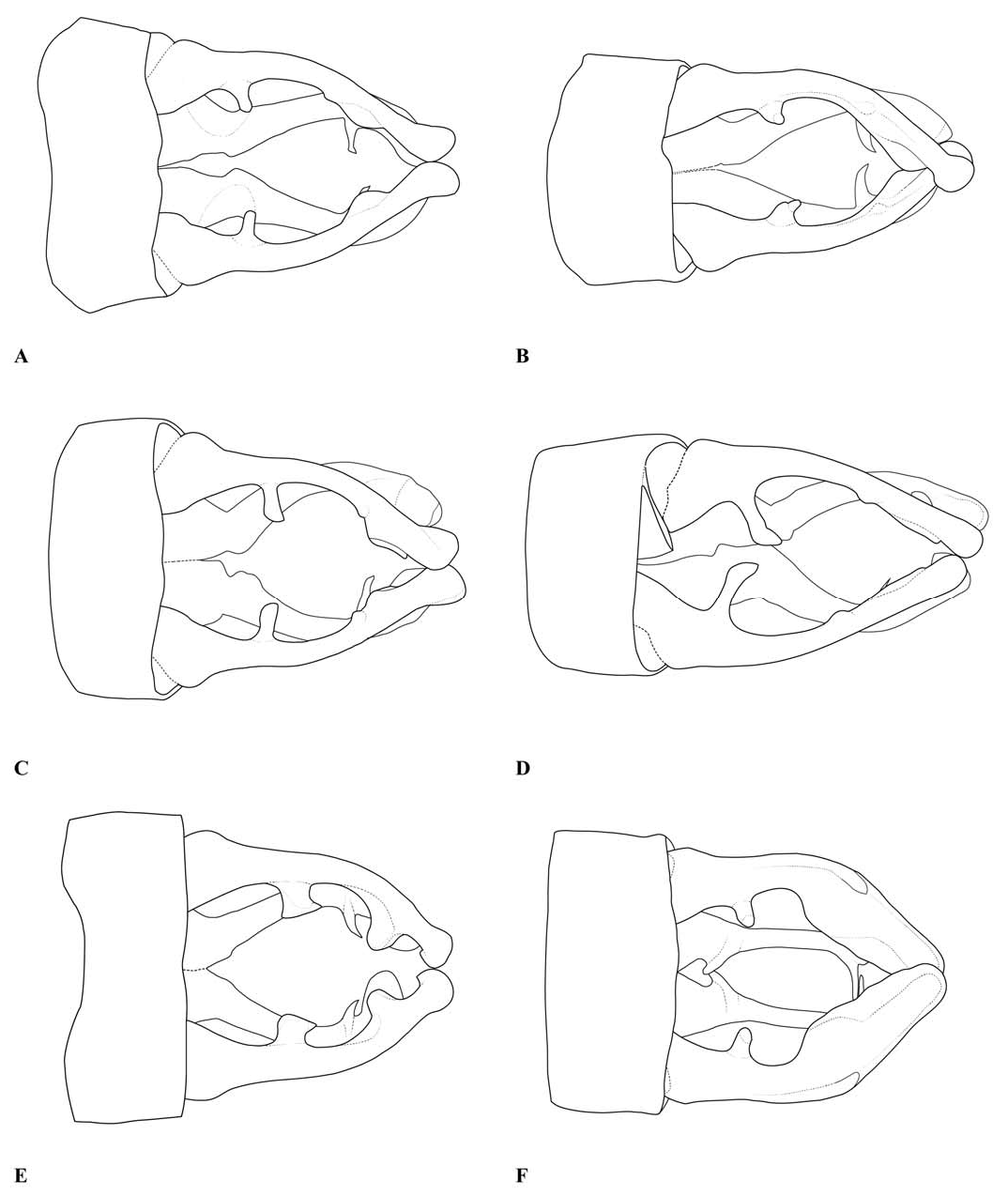
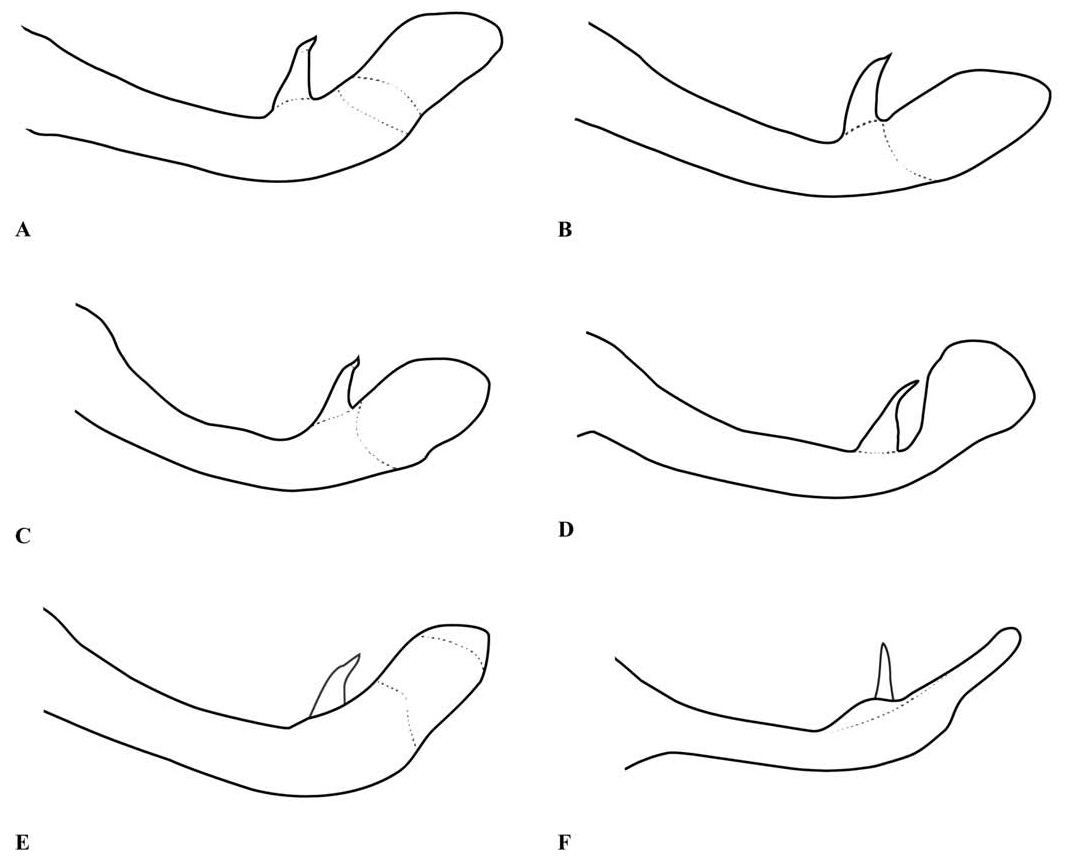
Abdomen: Slender, mostly dark dorsally with variably developed basal annuli on S2–7. Always with some pale markings on at least one of S8–9. Penis: with a row of prominent setae on shaft. Internal fold well developed. Terminal part with a pair of long distal horns, curved downwards and then upwards at tip; between these horns is a prominent convex tongue like structure (e.g., Fig. 1C View Fig ). Male anal appendages (e.g., Figs. 2 View Fig , 6–8 View Fig View Fig View Fig , 12–14 View Fig View Fig View Fig , 17C, D View Fig , 18A–D View Fig ) with short setae over much of their surfaces, not mentioned further unless of diagnostic importance and normally omitted from the illustrations. Superior appendage moderately broad at base, becoming narrower shortly afterwards, in lateral view with the upper margin running straight until turning down at one-half to two-thirds length; lower margin bent sharply upwards shortly after base, then running straight to the down-turn, giving an arched appearance to the whole. There is a structure visible in dorsal and sometimes lateral view, situated from one-quarter to slightly less than half the length in dorsal view, arising from the interior of the appendage, and referred to as the interior projection of the superior appendage hereafter. The interior projection is usually strongly developed, and variously shaped, from a mere bump or ridge to a simple spur or a blade like structure with dorsal and/or ventral spurs. At or around the level of the down-turn there is a dorsal interior structure, referred to as the dorsal projection of the superior appendage, usually poorly developed and visible as a slight bulge in dorsal view and sometimes as a distinct angled peak in lateral view, but usually best seen from a interior view. However the dorsal projection is better developed in a few species. The apical section from down-turn to tip is typically expanded and variously shaped. In T. tubau the interior projection is absent and the dorsal projection is just a slight bump placed somewhat interiorly and not visible in the illustrations in Dow (2010).
Inferior appendage broad at base but immediately tapering, with a complex membranous articulation close to the base, after which cylindrical (this part referred to here as the stem) until about the level of the down-turn of the superior appendage, where dorsal-ventrally flattened and expanded laterally to a greater or lesser degree in most species, this section usually hollowed dorsally to form a scoop-like structure, and referred to here as the scoop of the inferior appendage. The concave surface of the scoop typically has a clump of setae at its base (not shown in illustrations) and normally faces upwards, but in some species it is twisted so that it faces outward or inwards in its terminal part. A spine, directed inwardly and typically at least slightly upwards and rearwards, originates dorsal-interiorly behind the scoop, and is often continued dorsally as a ridge behind it.
Female anal appendages and ovipositor ( Fig. 1D View Fig ): Superior appendages shorter than S10, approximately triangular in profile. Inferior appendages short, rounded and unremarkable.
Ovipositor typically long, extending well beyond the tips of the superior anal appendages.
Remarks. — The nearest allies of Telosticta spp. appear to be three Bornean species: Drepanosticta actaeon Laidlaw , D. rufostigma (Selys) (which are clearly closely related to each other), and D. dulitensis Kimmins. None of these species bear the distinctive lateral processes on the pronotal hind lobe that are present in Telosticta , or a tongue-like structure between the horns of the terminal segment of the penis. D. actaeon and D. rufostigma also lack antehumeral marks and dorsal blue markings on the terminal abdominal segments; these characters are variably present in D. dulitensis . D. actaeon and D. dulitensis have a significantly wider vertex than Telosticta , but D. rufostigma is very variable in this character and overlaps the range seen in Telosticta . There are similarities in the general form of the anal appendages of Telosticta and D. actaeon , D. dulitensis , and D. rufostigma , but the scoop of the inferior appendage is modified into a strap like structure in these species (a condition approached in the inferior appendages of T. gading ). In D. dulitensis the interior projection of the superior appendage is present, in a basal position, and there is also a dorsal projection. In D. actaeon and D. rufostigma a dorsal spine is present on the superior appendage, in the position of the interior projection of Telosticta and D. dulitensis ; this is likely to be homologous with the interior projection of Telosticta . There is also a small dorsal projection close to the tip in D. actaeon and D. rufostigma . Orr (2003: 69) included D. dulitensis with the species here placed in Telosticta in his group 1 of the Bornean Platystictidae ; it certainly appears to be closest relative of Telosticta but is distinctive enough to merit a genus of its own which would be the sister of Telosticta . D. actaeon and D. rufostigma form a separate clade.
The lateral processes of the posterior pronotal lobe in Telosticta are almost unique in the Platystictidae . Some other species, for instance Drepanosticta sbong Dow and D. hamadryas Laidlaw and its allies from Peninsular Malaysia, have the lateral extremities of the posterior pronotal lobe produced into short downwards-directed processes. In D. sbong these processes are directed rearwards and slightly outwards, different from any species placed in Telosticta ; D. sbong differs considerably in many other characters and is not likely to be closely related to Telosticta . In D. hamadryas and its allies, the lateral processes are very small and it is questionable if they are homologous to those of Telosticta .
The tongue-like structure, or at least a slight convexity between the horns of the head of the penis, is present in some other species groups within the Sundaland clade, for instance in D. barbatula Lieftinck and its allies, where it is very poorly developed. Drepanosticta elongata Wilson & Reels, 2001 has a very long tongue-like structure in the same position as Telosticta (Wilson & Reels, 2001: Figs. 52, 53).
There are similarities in the anal appendages of Telosticta with not just D. actaeon , D. dulitensis , and D. rufostigma , but many other species occurring in Sundaland and further afield. Spines or tubercles likely to be homologous to the interior or dorsal projections of Telosticta are present on many species. The general plan of the inferior anal appendages, with an articulated basal area, narrow stem and sub apical dorsal-internal spine followed by a modified apical section is repeated in many species, including species from China ( D. elongata ) and the Philippines ( Drepanosticta lestoides (Brauer)) . This form of anal appendages differs from that in species such as Protosticta foersteri Laidlaw from Peninsular Malaysia and its allies from further north, or Drepanosticta forficula Kimmins from Borneo, or most species from the Philippines and further south and east.
In the molecular phylogenies of the Platystictidae presented in van Tol et al. (2009) the one species included that is here placed in Telosticta ( Protosticta aff. feronia , = T. dayak , new species) forms a monophyletic group with D. rufostigma and D. lestoides ; D. actaeon and D. dulitensis were not included in these results. It appears likely that Telosticta falls together with a number of other species in a larger clade, comprising a number of as yet unnamed genera. This group can, for convenience, be called the “Sundaland” clade, although it includes species occurring well beyond Sundaland, and certainly does not include every species occurring in Sundaland. The Sundaland clade can be characterised in terms of similarities in the structure of the anal appendages, and in turn falls within the West Malesian clade of van Tol et al. (2009). Conversely, P. foersteri and its allies fall within the West Malesian clade but not the Sundaland clade, and most Philippine species are outside the West Malesian clade.
Within Telosticta provisional species groups can be recognised, here named either after the first described species, or the most widespread species, in each group. The atypical T. tubau is considered to form a group of its own, characterised by S10 with long dorsal setae and superior anal appendages without interior projection. The dupophila - group consists of four species ( T. dupophila , T. gading , T. santubong , and T. serapi ) from western Borneo, all bearing short to moderate length lateral processes on the posterior pronotal lobe, vein ab typically present, superior anal appendages with a weakly developed dorsal projection and apical part expanded, flattened but not folded and without a notch or cleft in the tip, and inferior appendages with the stem relatively robust and the spine placed dorsally and directed strongly rearwards, so only partially or not at all visible in a ventral view. The feronia- group consists of species with moderate to long lateral processes on the posterior pronotal lobe, vein ab present or absent, superior anal appendage with a weakly developed dorsal projection and its apical part expanded, flattened but not strongly folded and with a small notch to a deep cleft in its tip. This group can be further divided into the feronia -subgroup ( T. bidayuh and T. feronia ) with a western distribution, moderate length lateral processes and only a small notch in the tip of the superior appendage, and the dayak -subgroup ( T. berawan , T. dayak , T. kajang , T. paruatia , and T. ulubaram ) with a more eastern distribution, long lateral processes and a conspicuous cleft in the tip of the superior appendage. The affinities of the remaining species are unclear, although T. longigaster may actually belong with the feronia -group and the remaining two species ( T. belalongensis and T. janeus ) may form a distinct group.
The anal appendages of Telosticta are of major diagnostic importance, but are structurally complex and must be examined very carefully when identifying species. In particular the shape of the terminal parts of both superior and inferior appendages can look very different depending on the angle from which they are viewed. The articulation of the inferior appendage is also complex, but does not appear to be of diagnostic value within the genus, and is frequently distorted or obscured; for this reason the illustrations give only a general impression of this structure.
Of the 15 named species included in the genus, 12 occur in Sarawak and Brunei. By comparison with Sarawak and Brunei, relatively little sampling of Odonata in habitats suitable for Telosticta has been conducted in Kalimantan and Sabah and more species from the genus can certainly be expected from these areas, and from Palawan.
The key below is organised for convenience of use rather than by species groups. Within the feronia -group, the feronia -subgroup and the dayak -subgroup appear to have allopatric distributions, but the feronia -subgroup species can occur sympatrically with species from the dupophila - group; for this reason all of the western species are treated together in the text following the key and in the illustrations. Similarly some species of the dayak -subgroup are known to occur allopatrically with T. longigaster , so these species are illustrated together. T. belalongensis also occurs allopatrically with T. longigaster , but is probably more closely related to T. janeus and is illustrated with that species.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.