
Squatina david, Arturo Acero P., José J. Tavera, Rafael Anguila & Luis Hernández, 2016
|
publication ID |
https://doi.org/ 10.1643/Ci-15-292 |
|
DOI |
https://doi.org/10.5281/zenodo.5629783 |
|
persistent identifier |
https://treatment.plazi.org/id/03DE663E-B828-AE4B-FF66-F981E443BAE2 |
|
treatment provided by |
Plazi |
|
scientific name |
Squatina david |
| status |
sp. nov. |
Squatina david View in CoL , new species
urn:lsid:zoobank.org:act:65AB97F7-86CA-47D6-898F- CA80A781E9B6
Figures 1–5
Squatina sp.—Uyeno and Sasaki in Uyeno et al. (1983): descriptive distinctive characters, distribution, remarks, and figures.–Paramo et al. (2015), ecologic and fishery information.
Squatina dumeril non Lesueur.—Valdez and Aguilera (1987), description, picture.–Cervigón (1992), description, figure.– Cervigón and Alcalá (1999), description, measurements, biological and ecological information, two pictures.
Squatina dumeril Lesueur. —Compagno (2002), map includes range of the new species.–Compagno et al. (2005), map includes range of the new species.–Robertson and Van Tassell (2015), map includes range of the new species.
Holotype.— INVEMAR PEC 8250 , 787 mm total length (TL), male, Colombia, Bocas de Ceniza , 11°0 0'33'' N, 75°0 3'19''W, 144 m depth, fishermen (gill nets), 12 July 2010. GoogleMaps
Paratypes.— INVEMAR PEC 8252 , 748 mm and 599 mm TL, two females, same locality and date as the holotype; GoogleMaps INVEMAR PEC 2186 , 257 mm TL, male, 11°0 5 '15 ''N, 74°36'27'' W, 100 m depth, 8 September 1996; GoogleMaps INVEMAR PEC 3988 , 245 mm TL, female, 12°30'33''N, 71°45'21''W, 150 m, 14 March 2001; GoogleMaps INVEMAR PEC 4117 , 302 mm TL, female, 11°23'0 7'' N, 74°0 8'41''W, 151 m, 19 March 2001. GoogleMaps
Diagnosis.— A Squatina distinct from other western North Atlantic squatinids based on the following combination of characters: mid-dorsal line lacking thorns or enlarged, sharp, pointed dermal denticles; eye-spiracle distance larger than 1.5 times eye diameter in specimens larger than 60 cm TL; each nasal flap with two rod-like barbels, outer barbel divided; coloration grayish to brownish yellow, male dark-spotted, females with abundant whitish spots.
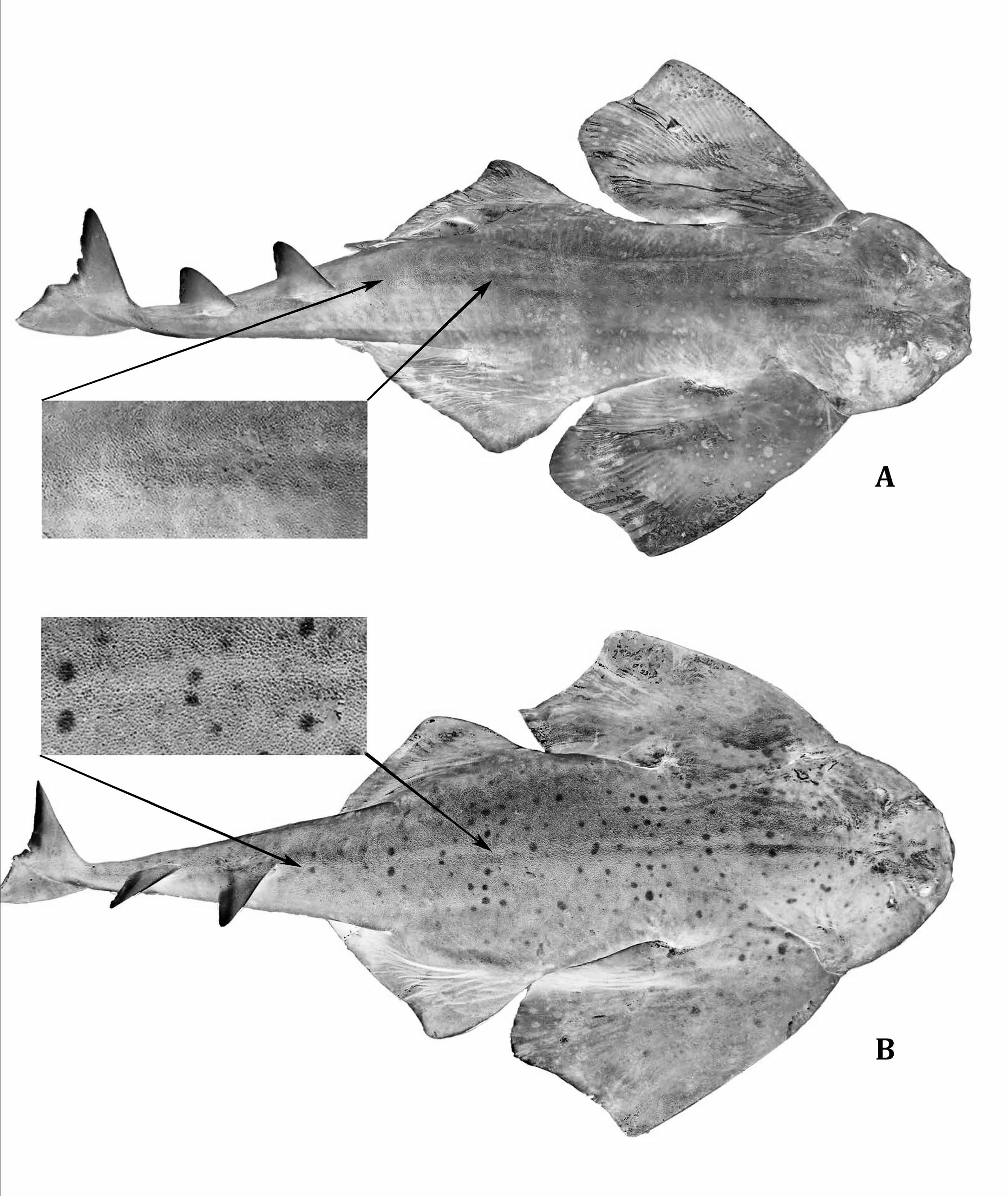
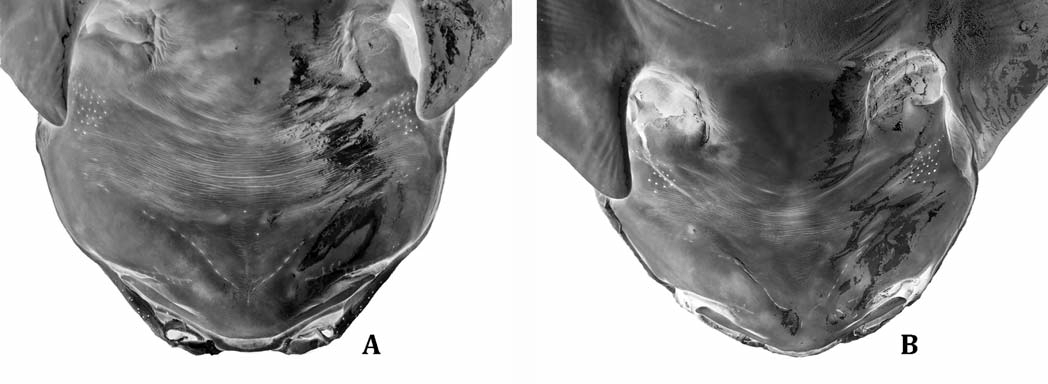
Description.— Dorsal surface covered with interspersed denticles of relative roughness, a densely concentrated patch of small curved spines along anterior pectoral-fin margins in mature males ( Fig. 1 View Fig. 1 A), poorly developed in females ( Fig. 1 View Fig. 1 B); ventral surface comparatively smooth, except for narrow bands of denticles along anterior margins of both pectoral and pelvic fins. Head rounded, length around five times in total length, with a maximum width occurring anterior of gill openings ( Fig. 2 View Fig. 2 ). Moderate tubercles interspersed above mouth and eye crests, smooth oval patch above midpoint of mouth in between eyes. Eyes almond-shaped, closely set, with an interorbital space 11– 13 times in TL; eye-spiracle distance relatively large, 30–43 times in TL. Spiracles crescent-shaped with pronounced interior papillae along posterior margin. Interorbital space around 1.1 times larger than interspiracle space. Center of upper lip arch exposed at midpoint of upper jaw, exposure semi-oval in shape. Labial furrows, roughly equal in length, extending from corners of mouth medially, with upper labial furrow partially covered with dermal folds ( Fig. 3 View Fig. 3 ). Distinct nasal flaps protruding from dermal folds above mouth, two barbels protruding from each flap; inner nasal barbels rod-like, containing little if any fringe; outer nasal barbels rod-like with a small branch protruding ventrally near tip ( Fig. 4 View Fig. 4 ). Nostrils large, protruding slightly, and tear-shaped. Dermal folds along exterior of head, one small lobe present at corners of mouth extending ventrally. Mouth length about 0.3 times as long as mouth width. Dentition consisting of small, dagger-like teeth, conical without cusplets on a broad base, in two orderly longitudinal rows on upper jaw, three rows on bottom jaw, no teeth at symphysis, 10 teeth by row in each jaw.
Pectoral fins large and broadly rounded, originating just behind gills. Anterior margin of pectoral fin mostly straight, 0.8–0.9 as long as pectoral length, extending to a lateral apex. Margin from lateral apex to posterior-most lobe slightly concave. Posterior lobe broadly rounded. Pectoral inner margin about one half of pectoral length, convex, and with a small lobe near pectoral base.
Overall pelvic fin shape somewhat triangular with rounded fin tips. Pelvic fins originating anterior to pectoral fin free rear tips. Pelvic fin length between one half and four fifths as long as pectoral fin length. Pelvic-fin base usually longer than pectoral-fin base, anterior margin 0.5–0.65 times as long as pelvic fin length. Posterior margin of pelvic fin straight to posterior free tip approximately 0.7 times fin length. Pelvic inner margin straight and short, approximately 0.4 times as long as pectoral fin length. Pelvic-fin insertion furrows on ventral side extend in a narrow ellipse to anterior apogee of vent, vent is within ellipse. One pelvic-fin tip extends to first dorsal origin, the other does not.
Dorsal fins slightly angular, second dorsal fin slightly smaller than first dorsal fin, with denticles covering the entirety of both fins ( Fig. 5 View Fig. 5 ). Dorsal-caudal space 1.0–1.4 times longer than interdorsal space. Anterior margin of both dorsal fins straight, nearly equidistant. Dorsal-fin bases subequal. Posterior margins straight, about 0.5–0.7 times as long as anterior margins. Inner margin of dorsal fins straight, approximately 0.2–0.5 times as long as anterior margins.
Caudal peduncle compressed dorso-ventrally with lateral longitudinal ridges, tapering posteriorly. Caudal fin lobelike, markedly at dorsal apex, dorsal margin broadly rounded, about 0.7–0.9 times as long as preventral caudalfin margin. Subterminal caudal-fin margin approximately 1.4 times longer than caudal upper postventral margin. Caudal lower postventral margin slightly convex, approximately 1.5 times longer than caudal upper postventral margin.
Coloration.— Grayish to brownish yellow. Male with dark spots, subequal to smaller than eye diameter ( Fig. 1 View Fig. 1 A). Females with abundant whitish spots, smaller than eye diameter ( Fig. 1 View Fig. 1 B).
Distribution.— Based on the type material, it is known from the Colombian Caribbean, between Bocas de Ceniza (11°0 0'33 ''N, 75°0 3'19''W) and Punta Gallinas (12°30'33''N, 71°45'21'' W). However, according to descriptions and figures of Uyeno et al. (1983) and Cervigón and Alcalá (1999), the species reaches at least to Suriname. Therefore, its distribution goes from Colombia to Suriname along the northern coast of South America.
Remarks.— Although similar to other northwestern Atlantic angel sharks, S. david can be distinguished from the other three species known from the area as follows. Its eye-spiracle distance in eye diameter is 1.5–2.1 times in specimens larger than 599 mm TL (0.8–1.3 in those 245–302 mm), while in S. dumeril is always less than 1.5 (Compagno et al., 2005). This ratio may be also larger than 1.5 in the two recently described Gulf of Mexico species (Castro-Aguirre et al., 2006). Also, its mid-dorsal line lacks thorns or enlarged, sharp, pointed dermal denticles, as in S. heteroptera and S. mexicana , while S. dumeril has a mid-dorsal row of hooked thorns.
Colonello et al. (2007) suggested the occurrence of pectoral spines on mature males, which might be widespread within the order Squatiniformes . The only adult male available (787 mm TL) of S. david indeed has a patch of spines on its pectoral fin edges, a trait associated to sexual dimorphism. In the numerous previously examined (Bigelow and Schroeder, 1948; Colonello et al., 2007; Baremore, 2010) morphometric differences between males and females, no mention has been made of nostril width. This tear-shaped cavity is wider, almost two-fold in adult males of S. david compared to females. We speculate that males may have a larger nostril area enhancing their capability to detect female reproductive
cues as has already been suggested by Arif (2011) for Scyliorhinus canicula .
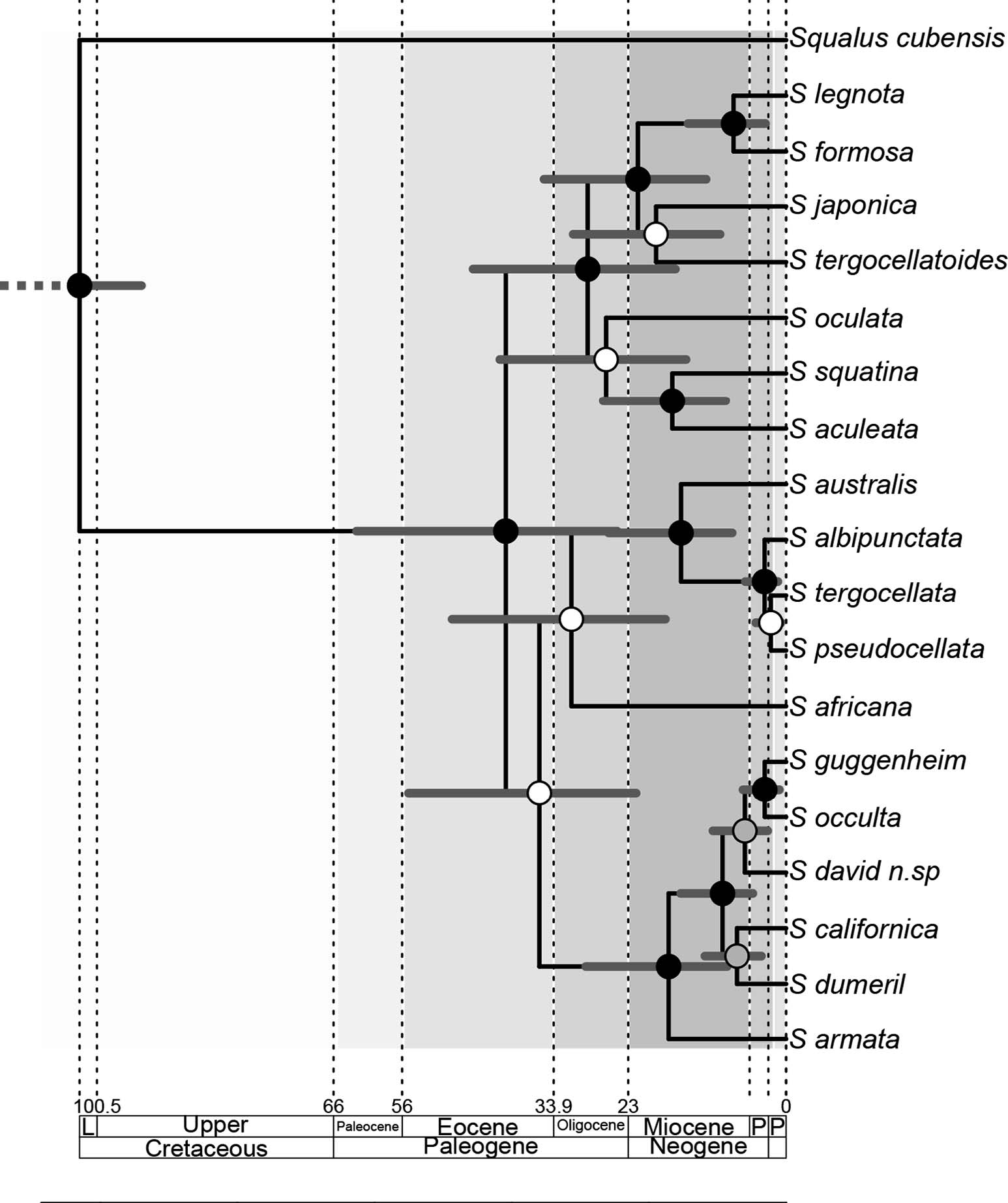
Stelbrink et al. (2010) included in a comprehensive worldwide species coverage, corresponding to world major geographic regions, four recovered Squatina clades. After including sequences from S. david , all reconstructions yielded the same results, with the new species being nested inside the American branch. All phylogenetic methods used in this study consistently recover S. david as the sister species of the branch, including the Brazilian S. guggenheim and S. occulta . According to our time-calibrated phylogeny, the branch including all South and North American species originated at some point between mid-Oligocene to late-Miocene (29.3– 8.6 mya). Our result displaces the earliest boundary of such splitting several million years before the hypothesis of Stelbrink et al. (2010). In accordance with our results, the 95 HPD credible interval of the time of origin of S. david spans from 10.7 to 2.7 mya, somewhere between mid- Miocene to Pliocene ( Fig. 6 View Fig. 6 ).
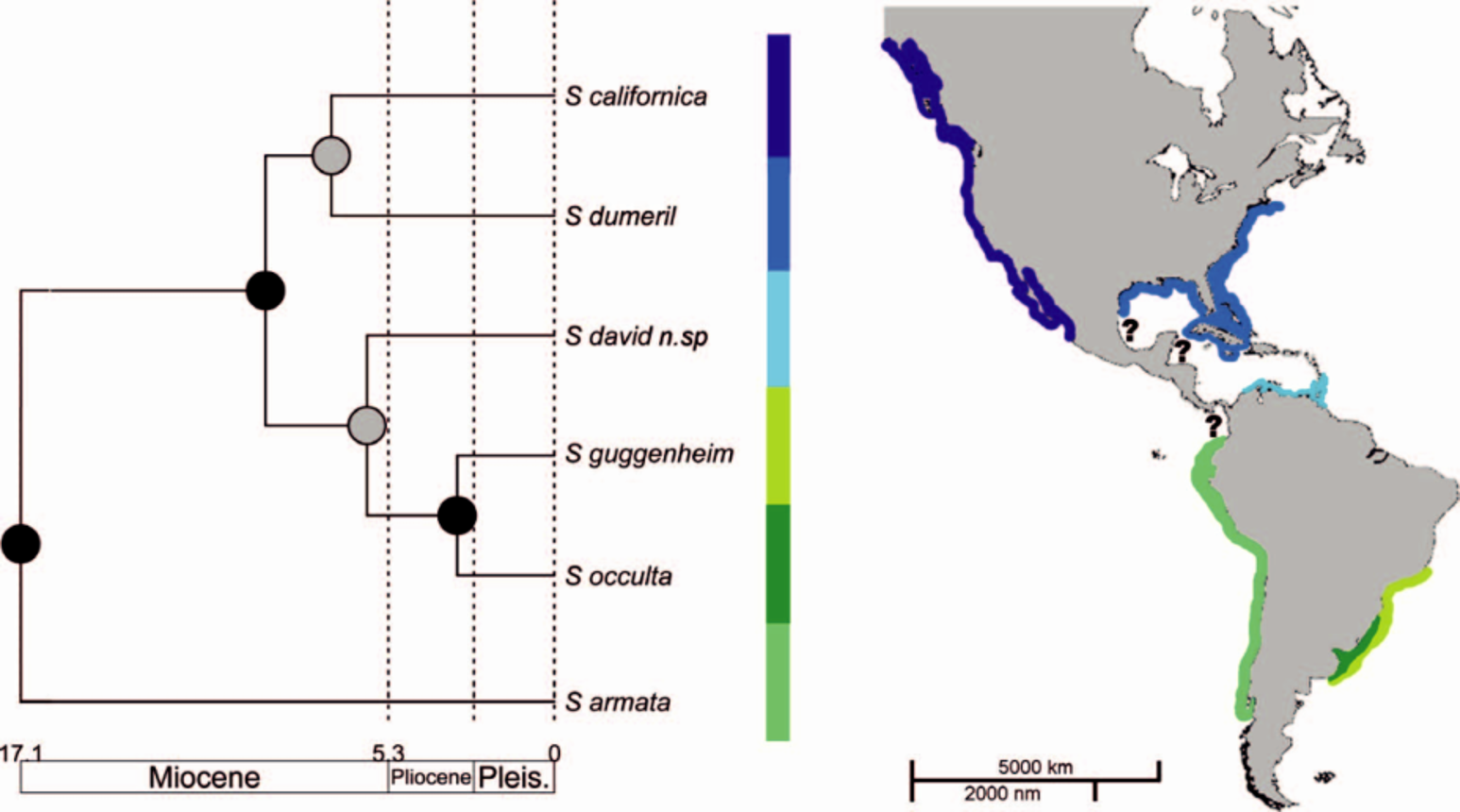
The branching arrangement of American Squatina exhibits a distinguishable biogeographic pattern, with the most basal species being restricted to the southeastern Pacific, one clade exclusive to South America, and a third being totally North American ( Fig. 7 View Fig. 7 ). Unfortunately, there are no available molecular data for the two endemic Gulf of Mexico species ( S. mexicana and S. heteroptera ), information that eventually may concur with the south-north grouping hypothesis. Geologic and paleoclimatic events during the Miocene-Pliocene associated with the rising of the Central American isthmus and its collision with northwestern South America (Coates and Obando, 1996; Coates et al., 2004) could have deeply influenced the evolutionary history of American angel sharks. A thorough review including the two Gulf of Mexico species and additional genes are necessary in order to have a better understanding of the evolutionary history of this American clade of elasmobranchs.
Etymology.— To be used as a noun in apposition. To honor the son of the first author, David Acero, who passed away on March 7, 2011. Pronunciation of david : dahveed.
Common names.— The following common names are proposed for the new species: David’s Angel Shark (English) and angelote de David (Spanish) .



| INVEMAR |
Instituto de Investigaciones Marinas de Punta de Betin |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |