
Rotundopteryx notata ( Duckhouse, 1966 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4457.2.4 |
|
publication LSID |
lsid:zoobank.org:pub:486B0557-DA8D-4042-A69B-1AAAEFFFE915 |
|
DOI |
https://doi.org/10.5281/zenodo.5975760 |
|
persistent identifier |
https://treatment.plazi.org/id/03DE8784-FFC7-105D-29B0-FC8BFA5018F5 |
|
treatment provided by |
Plazi |
|
scientific name |
Rotundopteryx notata ( Duckhouse, 1966 ) |
| status |
|
Rotundopteryx notata ( Duckhouse, 1966)
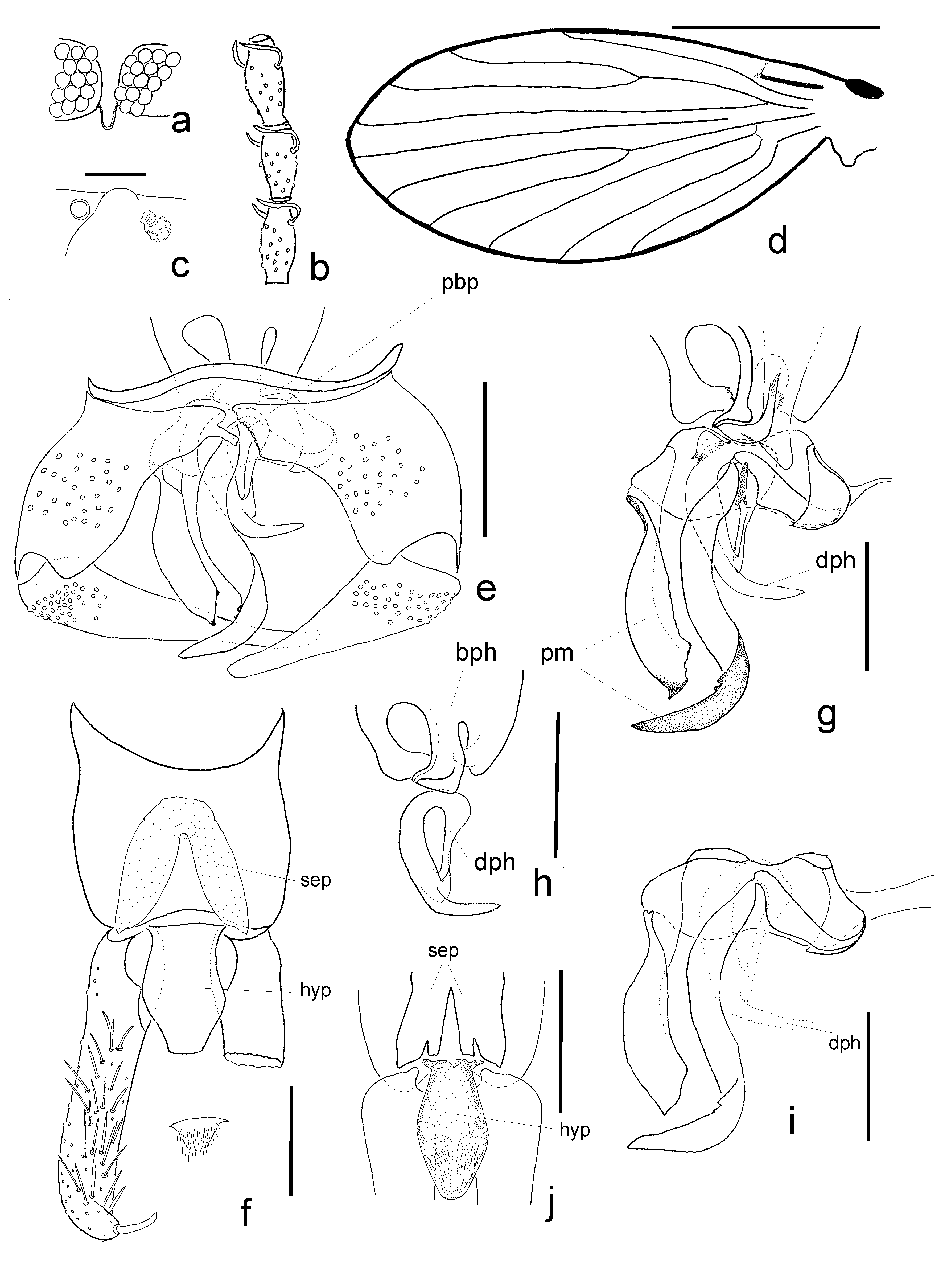
( Figs 4 a–i View FIGURE 4 )
Material. 1♂ New South Wales, sidestream of Wollondilly river near Goodmans Ford, 20–21 September 1980, leg. Theischinger. Distribution : Australia: Australian Capital Territory, first record for New South Wales .
Redescription. Head with eye bridge of 5 facet rows, interocular suture narrow, deeply U-shaped, distance between the eyes 1 facet diameter; 1 row of postocular bristles; apical end of head incurved; clypeus and frons separated; antenna with scape, pedicel and 14 flagellomeres; scape tubular, longer than wide, pedicel spherical; flagellomeres barrel-shaped, flagellomeres 2–12 with a pair of short digitate ascoids, on first and apical 2 flagellomeres not undoubtedly discernable; apical flagellomere with apiculus. Relative length of antennal articles: 28-26-22-22-22-22-19-19-18-17-17- 16-16-13-10 -8; absolute length: 0.07-0.07-0.06-0.06-0.06-0.06-0.05-0.05- 0.05-0.04-0.04-0.04-0.04-0.03-0.03- 0.02 mm. Four palpus segments, terminal segment flexible; relative length: 45-68-66-92; absolute length: 0.12-0.17- 0.17-0.24 mm.
Thorax with scent organ in frontal section of mesothorax below stigma; it is probably extendable with a spherical head with several circular apertures; diameter of head of the size of stigma.
Wing length 2.50 mm, width 1.10 mm; length by width index 2.27; Sc terminates in wing, faint crossveins towards C and R1; radial and medial forks about middle of wing, tip of Cu basal of forks; crossveins r4-r5 and r5-m1+2 faint. CuA long almost straight, CuP short straight faint.
Abdomen consists of 8 segments and inverted genitalia, gonocoxites and aedeagus asymmetrical. Hypandrium is an equally thin, slightly curved band. Goncoxites asymmetrical, tube shaped, distally narrowed, basal width almost twice the distal width; left gonocoxite with a short parabasal process (sensu Duckhouse 1990); its counterpart without such process but with serrate inner margin. Gonostyli approximately 1.8 times as long as gonocoxite, almost straight, distally narrowed. Parameres asymmetrical twisted bands shifted to the side; the one near the gonocoxite with parabasal process short broad; the other one longer and thinner with the tip bent at right angle in the direction of its counterpart. Both evolve from an asymmetric parameral bridge between the gonocoxites; the bridge is connected by straight sclerite to the gonocoxite, this part was not observed on the other side. Basiphallus sclerite very broad almost parallel sided; distal end with three lobes, lateral ones asymmetric, turned towards the undulating front edge of the parameral bridge; the median process is bent towards the distiphallus. Distiphallus sclerite is an asymmetric triangular ring shaped sclerite with a long apical spur pointing sideward. Epandrium rectangular, anteriorly curved, with a single foramen. Surstyli almost straight, only distal end curved, about twice the length of the epandrium, with 1 large apical retinaculum and 20–25 accessory retinacula along its length on ventral side. Subepandrial plate V-shaped, both lobes apically split so that lateral broader parts are jointed with the surstyli, the inner narrower parts are jointed to the well sclerotized basis of the hypoproct. Proctiger with a large cone shaped hypoproct and a small setose epiproct.
Comments. Duckhouse (1990) created the genus Rotundopteryx for most Australian and Tasmanian species that until then were placed in Pericoma Walker (type-species: R. sylviae Duckhouse , orig. des.). The original description of the genus has to be supplemented by some features. Genitalia are remarkable in that most parts are asymmetrical; this includes gonocoxites, parameres, basiphallus and distiphallus. Gonocoxites are distinguished by the shape of basal inner margin (parabasal process, Duckhouse 1990) on the ventral side; the bridge between gonocoxites evolves from the dorsal edge of the gonocoxites, and from this bridge a pair of unequal sized and shaped parameres develops. The asymmetry of the gonocoxites is not as eye catching as in Neurosystasis species from Cuba, in that even the general shape of left and right gonocoxites is fundamentally different ( Wagner & Kvifte 2017). The lateral lobes of the basiphallus terminate close to the areas from which parameres descend; the inner lobe (broader, probably 2 lobes are very close to one another or even stacked?) approaches the distiphallus. The distal split of the subepandrial plate and the joints of the inner lobes with the hypoproct—clearly visible in R. coracina and R. satchelli —are as well remarkable. It is notable to mention that particular specimens of R. satchelli display laterally reversed aedeagi (mirror-image asymmetry, or antisymmetry) as is known from other psychodids with asymmetrical genitalia (e.g. Wagner 1979).
The present redescription of R. notata is not based on the holotype; however, genitalia are undoubtedly identical. Compared with the original description there are some differences in the relative length of the palpus and antennal segments and in the distance between the eyes that however fall in the range of variation of every species.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.