
Berrulestes phelizoni, Hooker & Russell, 2012
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2011.00787.x |
|
persistent identifier |
https://treatment.plazi.org/id/03DE8792-FF83-6558-FC7B-FE85FC2EFC58 |
|
treatment provided by |
Marcus |
|
scientific name |
Berrulestes phelizoni |
| status |
sp. nov. |
BERRULESTES PHELIZONI SP. NOV.
( FIGS 5B View Figure 5 , 8 View Figure 8 , 10, 11A, B, 14)
vp 1964 Pagonomus dionysi Russell ; Russell: p. 66, pl.
6, fig. 5b, 5e. vp 1967 Pagonomus dionysi Russell ; Russell, Louis &
Poirier: pl. 20, fig. 2 (top row middle, bottom row left).
Etymology: After Monsieur Alain Phélizon, for finding key specimens of the species, including the holotype dentary, and for his considerable aid to the authors based on his knowledge of Palaeogene faunas of Champagne and of the amateur palaeontologists who have worked there.
Holotype: R dentary with P 3 –M 3 and separate condyle, MNHN.F.BR-29-Ph, bed 5, Berru.
Paratypes: RP 4, MNHN.F.CR-1693-Pn; four LM 1 s, MNHN.F.CR-213, CR-1230, CR-59- V, CR-20- MD; six RM 1 s, MNHN.F.CR-210, CR-937, CR-5487, CR-1390-Pn, CR-1699-Pn, UCMP.62006(CR-4419); RM 2 , MNHN.F.CR-3-Bn; LP 4 , MNHN.F.CR-34- MD; RP 4 , MNHN.F.CR-1448-Pn; three RM 1 s, MNHN.F. CR-42-Pn, CR-1415-Pn, CR-1707-Pn; three LM 2 s, MNHN.F.CR-281-Bn, CR-333-Pn, CR-17477; three RM 2 s, MNHN.F.CR-11901, CR-307-L, CR-94-Pn; five RM 3 s, MNHN.F.CR-341-Pn, CR-1730-Pn, CR-329-L, CR-330-L, CR-56- MD; RDP 4 , FSL.1.881; all Cernay .
LP 4, MNHN.F.BRL-79-P; RM 1, MNHN.F.BRL- 203-P; L dentary with M 2–3, MNHN.F.BRL-7-F; RM 3, MNHN.F.BRL-18-G; all bed 4, Berru.
RM 1, MNHN.F.BR-6-L; LM 2, MNHN.F.I-283; RP 4, MNHN.F.I-602; RM 1, MNHN.F.I-710; LM 3, MNHN.F.BR-125-L; three RM 3 s, MNHN.F.I-580, I-754, I-247; all bed 5, Berru.
LM 3, MNHN.F.R-201-Lass, Cernay or Berru.
Referred specimens: LP 4, MNHN.F.CR-17476; RP 4, MNHN.F.CR-237-Bn; 2 LM 1 s, MNHN.F.CR-384, CR-282-Bn; LM 3, MNHN.F.CR-1424-Pn; all from Cernay.
RM 1, MNHN.F.I-606; LM 2, MNHN.F.I-692; RM 2, MNHN.F.I-689; all bed 5, Berru.
Doubtfully referred specimen: LDP 4, MNHN.F.CR- 374; Cernay.
Age and distribution: Sables de Châlons-sur-Vesle supérieurs, late Thanetian, Late Palaeocene, Cernay and Berru (beds 4, 5), France.
Diagnosis: Medium-sized species of Berrulestes , mean length of M 1 3.12 mm. Cusps of cheek teeth somewhat bulbous. P 4 metaconid extending mesially (shared with B. pellouini ). M 1–2 with deep, acute postflexi. M 1 hypocone subequal in height with protocone (shared with B. pellouini ). M 1 postcingulum not joining confluent postmetaconule crista and metacingulum (shared with B. poirieri ). Lower molars with paraconid cuspate, small and low, less than half the height of the metaconid on M 1 in lingual view. Lower molar precingulid shelf-like, bulging mesially (shared with B. pellouini ). M 1 talonid of equal length and width to trigonid. M 1–2 protoconid height equal to or less than tooth width. Dentary deepening posteriorly.
Description
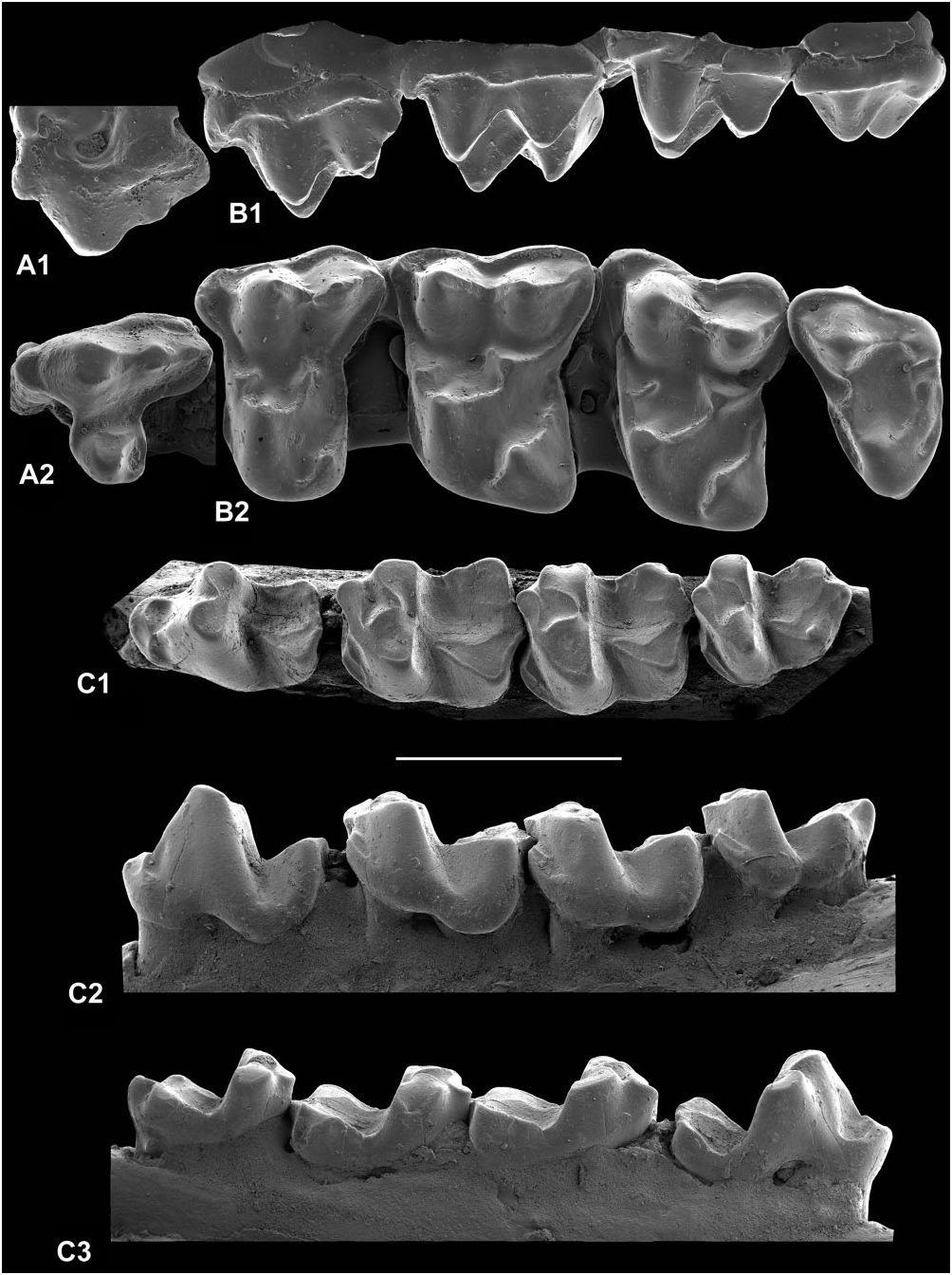
P 4: In both specimens the metacone is smaller than the paracone (Fig. 10A1), but the difference is not as great as in W. girardi . The parastyle is large and mesially projecting (Fig. 10A). Although in one it is broken, what remains suggests one even larger than that of the complete specimen. The outline is ‘waisted’, there being a distinct postflexus and a slight preflexus enhanced by the large parastyle. The postcingulum is weak and on it rests a very small hypocone. In one specimen the postprotocrista joins the lingual end of the metacingulum, whereas in the other the postprotocrista ends before reaching the metacingulum. In this specimen the metacingulum has a small cusp developed at its lingual end. The postflexus is also shallower. Length is 2.84 mm, width is 2.96–3.15 mm.
M 1: The cusps are rather bulbous, matching the lowers in this respect. The absence of a connection between the postcingulum and postmetaconule crista–metacingulum (Fig. 10B2) is a consistent character, which differentiates B. phelizoni from the morphologically similar B. pellouini . It also enhances the depth of the postflexus in B. phelizoni . The precingulum is consistently present but varies in strength. There is also variation in development of the paraconule, varying from a distinct cusp to a slight swelling on the preprotocrista–paracingulum. The parastyle is prominent mesially (Fig. 10B). The presence of a short premetaconule crista is fairly consistent, although missing in one specimen. One specimen is unusual in having shallow pre- and postflexi, a relatively weak precingulum, and a hypocone that projects markedly lingually.
·
Figure 10. Scanning electron micrographs of gold-palladium coated epoxy casts of teeth and jaw of Berrulestes phelizoni gen. et sp. nov., late Thanetian, France. A, RP 4 (reversed), Cernay (MNHN.F.CR-1693-Pn). B, RM 1 (reversed), Cernay (MNHN.F.CR-937). C, LM 2, bed 5, Berru (MNHN.F.I-283). D, LDP 4, Cernay (MNHN.F.CR-374). E, RDP 4 (reversed), Cernay (FSL.1.881). F, RM 1 (reversed), Cernay (MNHN.F.CR-42-Pn). G, RM 2 (reversed), Cernay (MNHN.F.CR- 11901). H, holotype right dentary with P 3 –M 3 (reversed) from bed 5, Berru (MNHN.F.BR-29-Ph). Views are: buccal (A1, B1, C1, D1, E2, F2, G1, H2), occlusal (A2, B2, C2, D2, E1, F2, G2, H1), and lingual (E3, F1, G3, H3). Scale bar = 2 mm.
M 2: The two specimens are broadly similar to M 1 in cusp and crest pattern and flexi, but the postcingulum and metacingulum are confluent and in one case the postmetaconule crista does not join the metacingulum (Fig. 10C). The outline is slightly distally tapering and more transversely elongate than in M 1. One tooth has a short premetaconule crista, the other lacks one. Both have poorly developed paraconules without postparaconule cristae. They are 2.41– 2.01 mm long by 3.40–3.70 mm wide.
P 3: This tooth has only been recognized in the holotype dentary (Fig. 10H). It is 2.95 mm long by 1.32 mm wide. The tooth lacks a metaconid, its place taken by a crest running down the distolingual margin of the protoconid. There is a small cuspate paraconid. The single-cusped talonid has hardly any basin. There is strong shearing wear buccally for the paracone of a P 3. The latter presumably would have had only a tiny protocone with no substantial P 3 talonid basin with which to occlude.
P 4: The typical crestiform paraconid that is confluent with a prominent precingulid has a lingual basin and is separated from the paracristid, as diagnosed (Fig. 10H), but this structure is subject to some variation. One specimen with a weak precingulid has the paracristid confluent with the paraconid ( Fig. 11A View Figure 11 ). Another, which is corroded, has a small cuspate paraconid, apparently with no basin, but which is still connected to the precingulid and is separated from the paracristid. A third tooth has a taller more cuspate paraconid with only a small basin, whereas in a fourth the paraconid is again taller and cuspate, but also has a typical basin and separation from the paracristid ( Fig. 11B View Figure 11 ). There is also variation in the size of the entoconid ( Fig. 11A, B View Figure 11 ), although in most specimens it is much smaller than the hypoconid.
M 1: There is variation in the position of the attachment of the cristid obliqua to the back of the trigonid (Fig. 10F2, H1). Extremes are median versus distinctly buccal. There is also variation in the length of the precingulid, ranging from short in the holotype (Fig. 10H2) to extending round much of the protoconid. Buccal and lingual trigonid cusps may be occlusally divergent (until the top third of the protoconid) to convergent from near the base. The relatively low crown height (Fig. 10F1, F2) means that large areas of exposed dentine appear relatively early in wear (Fig. 10H). This is enhanced by the slightly bulbous cusps and the typically flat cusp-tip wear that dominates over buccal phase shear. It is instructive to compare the wear patterns of an unworn tooth with those of successive wear stages (Fig. 10F2, H1 for extremes).
M 2: These teeth have very small paraconids and the talonid is narrower than the trigonid. Otherwise they are similar to M 1 (Fig. 10G, H). There is variation in the extent of the precingulid and in trigonid width.
M 3: Like M 2, the paraconid is very small (Fig. 10H). The entoconid is small and crestiform in most individuals. It lies at about the same level as or slightly distal of the hypoconid. For size see Figure 14 View Figure 14 .
Deciduous premolars: CR-374 is a problematic upper molariform (Fig. 10D). It is 3.08 mm long by 3.02 mm wide, so is nearly equidimensional. It has equal-sized paracone and metacone, moderate enamel thickness, strong paraconule and metaconule that are largely devoid of cristae except for the preparaconule crista, tall hypocone that is slightly lower than the protocone, and buccal and lingual cusps that converge at an acute angle. Most of these features suggest a DP 4, according to their expression in M 1, although the enamel thickness is comparable to a permanent molar of Berrulestes or to a DP 4 of Gigarton meyeri , which has quite different morphology ( Fig. 17N). Size is suggestive of B. phelizoni , but certain features are atypical: the lack of a postmetaconule crista, a postcingulum that is confluent with the metacingulum, and a wide shallow postflexus. However, given that deciduous teeth may be rather variable, referral to B. phelizoni seems most likely given the available evidence.
FSL.1.881 is an elongate lower molariform, 2.80 mm long by 1.65 mm wide at the talonid. It is low crowned with a talonid wider than the trigonid (Fig. 10E). The protoconid and metaconid are slightly taller than the hypoconid and entoconid, but the paraconid is very small and isolated from the paracristid. The trigonid is thus open mesially and what remains of its basin is filled by a protoconid rib that extends nearly as far as the trigonid notch. There is a precingulid that extends distally right round the protoconid. The cristid obliqua is strong and joins the base of the trigonid back wall in a buccal position. It is thus judged to be a DP 4 of B. phelizoni . It differs from M 1 of this species only in being slightly smaller, more elongate, having a longer more open trigonid, a deeper protocristid notch, and slightly thinner enamel. It differs from DP 4 of Pa. dolloi and Pa. cf. dolloi in lacking a large mesially protruding paraconid ( Teilhard de Chardin, 1927: pl. 2, figs 22, 23; Godinot, 1981: fig. 21). This mirrors the small size and low height of the B. phelizoni P 4 paraconid (Figs 10H, 11A, B).
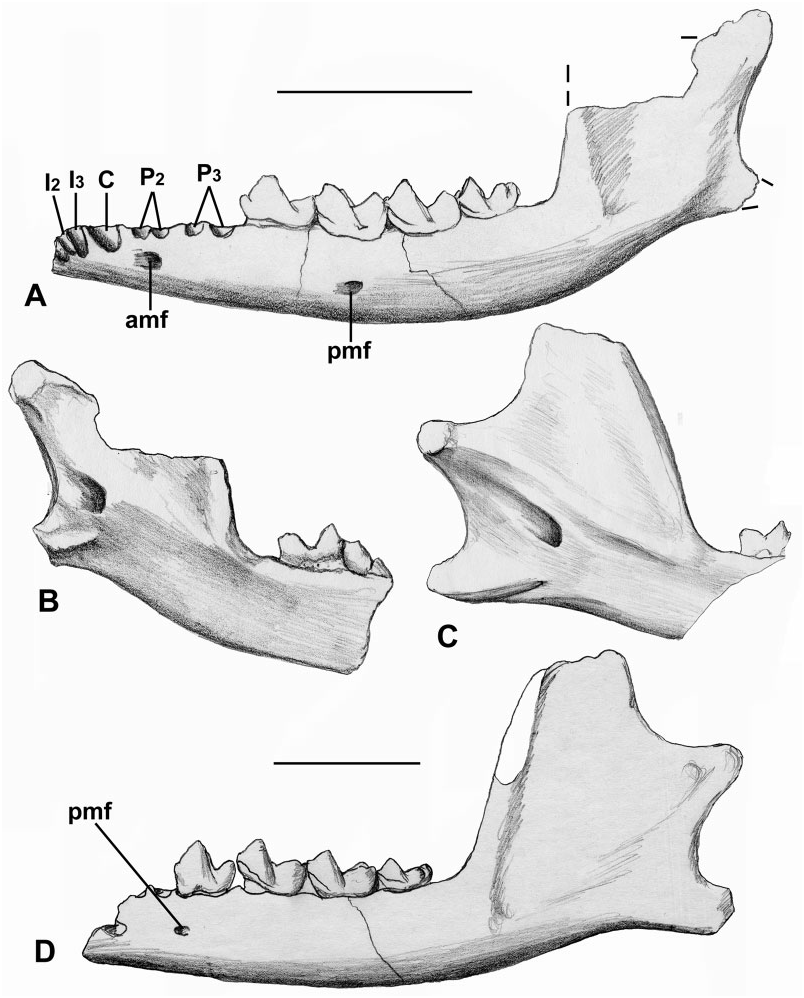
Dentary: The holotype dentary is in several nonfitting pieces ( Fig. 5B View Figure 5 ). It has P 3 –M 3 with crowns. The horizontal ramus deepens posteriorly, reaching its maximum a short distance behind M 3. It is incomplete posteriorly and lacks most of the ascending ramus. The posterior mental foramen is at the junction of P 4 and M 1 ( Fig. 5B View Figure 5 1 View Figure 1 ). In front of P 3, there is a short diastema, then two similar-sized circular alveoli, judged to belong to P 2. Part of the anterior mental foramen lies below its anterior alveolus. More anterior again there is the back of an apparently slightly elongate alveolus, belonging to P 1, which may therefore be single rooted ( Fig. 5B View Figure 5 3 View Figure 3 ). To be double rooted, one would expect the posterior root to be smaller than those of P 2 and circular. A more anterior fragment of dentary does not quite fit the main piece. It bears part of the symphyseal surface ( Fig. 5B View Figure 5 2 View Figure 2 ) and parts of four alveoli that lie obliquely to the dorsal surface, indicating procumbency ( Fig. 5B View Figure 5 1–3 View Figure 1 View Figure 2 View Figure 3 ). The most distal is the largest, but retains only its mesiolingual part. It is judged to belong to the canine. The next two more mesial alveoli are smaller and represent I 2–3. The mesial-most alveolus has only its distal wall preserved, but was apparently the same size as the succeeding two and is identified as I 1 ( Fig. 5B View Figure 5 2 View Figure 2 ). The condyle is preserved isolated. Its articular surface is very gently convex dorsally in posterior view ( Fig. 5B View Figure 5 8 View Figure 8 ) and more strongly convex in medial or lateral view ( Fig. 5B View Figure 5 4 View Figure 4 , B 5 View Figure 5 ). It projects slightly on the medial side, curving slightly ventrally. In dorsal view it is slightly broader than long, the outline tapering slightly medially ( Fig. 5B View Figure 5 6 View Figure 6 ). It is unlike the amphilemurid Gesneropithex in lacking a ventrally projecting posteromedial articular lobe; Leptictis and modern lipotyphlans differ in much the same way ( Butler, 1988: fig. 5.2). Modern macroscelideans are similar to B. phelizoni in lacking the ventrally projecting posteromedial lobe. Elephantulus and Rhynchocyon differ, however, in having more cylindrical, mediolaterally elongate condyles and in having the dorsal outline tapering laterally, not medially. Macroscelides proboscideus is the taxon most like B. phelizoni and differs only in having a gently concave dorsal surface in posterior view and a slightly greater ventromedial projection.
Discussion
In terms of length versus width measurements, those M 1 s and M 2 s belonging to Berrulestes from Cernay and Berru are significantly larger than those of either W. girardi or Pa. dolloi . They form three distinct clusters, although in the case of M 1 the largest ‘cluster’ consists of one plot ( Fig. 8A, B View Figure 8 ). In both cases the measurements of the holotype of B. phelizoni plot in the middle cluster. The coefficients of variation of the middle clusters ( Table 4) are within the range of variation expected of a single species ( Gingerich, 1974). If the measurements of each of these middle clusters are added to each of the respective clusters of smaller or larger measurements, the coefficients of variation increase significantly. They approximately double with addition of the smaller cluster or with the addition of the widths of the larger cluster ( Table 4), indicating three species. Thus the clusters of smaller lower molars are named as B. pellouini sp. nov. (q.v.) and the larger lower molars as B. poirieri sp. nov. (q.v.).
The situation is more complicated for the only other adequately represented tooth type, M 1, as there is only one cluster of measurements larger than Pa. dolloi , with single outliers of smaller and larger plots ( Fig. 8D View Figure 8 ). As for M 1 and M 2, length and width measurements of M 1 ( Table 4) have low coefficients of variation indicative of a single species. The length span overlaps extensively with that of B. phelizoni M 1, with a slightly smaller mean, so this M 1 cluster is also referred to B. phelizoni . Adding the small value outlying plot to this cluster increases the coefficient of variation somewhat ( Table 4), suggesting more than one species, so this small M 1 is referred to B. pellouini sp. nov. Adding the larger value outlying plot to the original cluster results in an even larger coefficient of variation, suggesting that the very large M 1 belongs to a species larger than B. phelizoni , namely B. poirieri sp. nov.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.