
Gibellula aurea, Mendes-Pereira & Araújo & Mendes & Fonseca & Alves & Sobczak & Góes-Neto, 2022
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.573.1.5 |
|
DOI |
https://doi.org/10.5281/zenodo.7334662 |
|
persistent identifier |
https://treatment.plazi.org/id/03DF2749-DE74-D853-638E-F853F346D580 |
|
treatment provided by |
Plazi |
|
scientific name |
Gibellula aurea |
| status |
sp. nov. |
Gibellula aurea View in CoL sp. nov. T. Mendes-Pereira, J.P.M. Araújo & A. Góes-Neto, Figs. 1 View FIGURE 1 and 2 View FIGURE 2 .
Mycobank number: —842796
Type: — BRAZIL. Ceará: Pacoti, Sítio São Luís, Trilha do Purgatório, 910 m elevation, 4°13’21” S, 38°53’35” W, 21 December 2019, collected by: Alves, J. E. R., on spider Macrophyes pacoti (Anyphaenidae) , deposited at the collection Herbário da UFMG–Fanerógamas, algas e fungos (BHCB); holotype: BHCB205962 , paratypes: BHCB205963; BHCB205964; BHCB205966 GoogleMaps .
Etymology: —from Latin, aurea : adjective of aurum, in allusion to the colour of gold. Named after the goldenyellow colour of the subiculum, which is characteristic of this species.
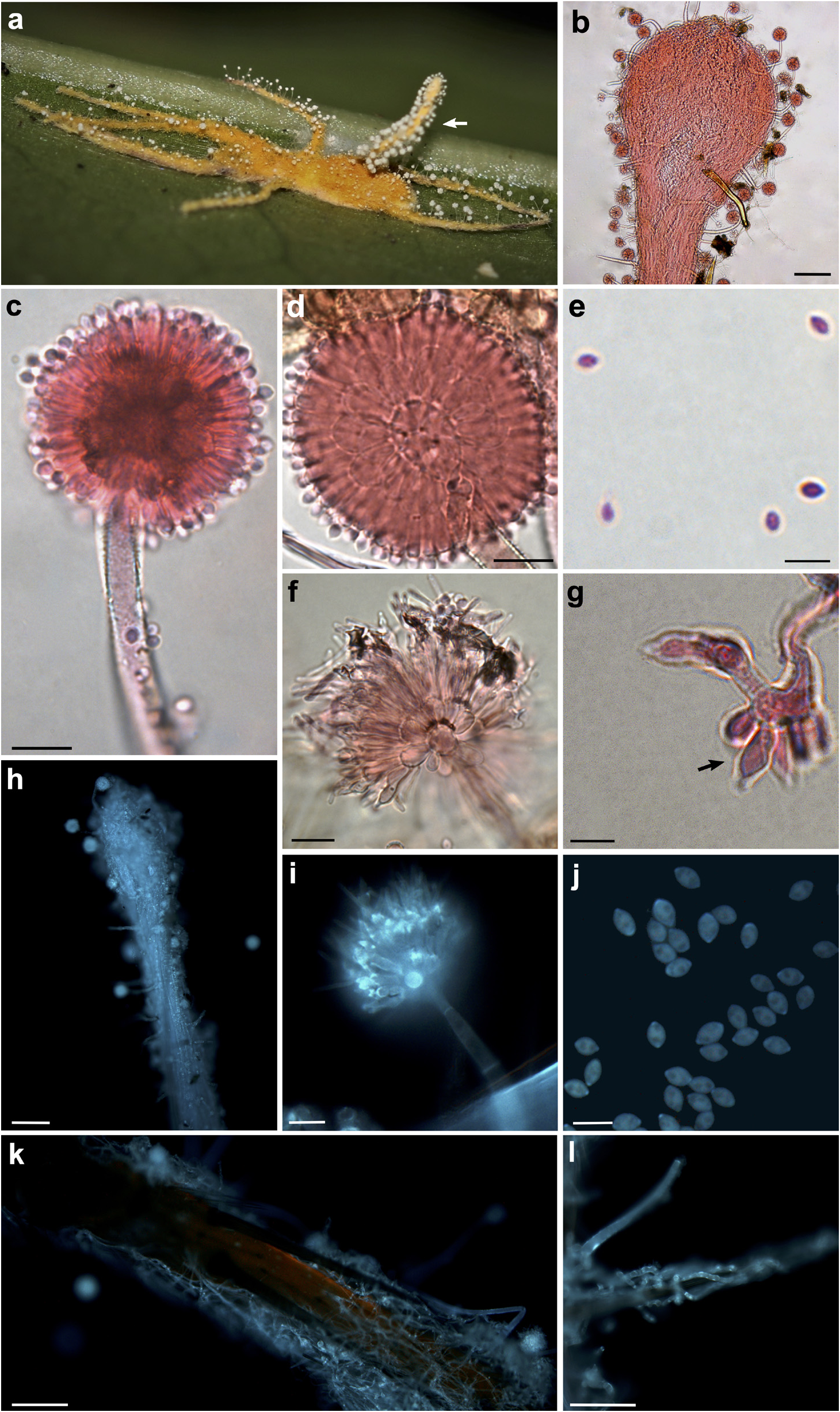
Diagnosis: — Gibellula aurea exhibits typical yellow hyphae, with white conidiophores ( Fig. 1a View FIGURE 1 ), differing from other species previously described in the genus.
Habitat: —Brazilian Atlantic Rainforest, Ceará State, Brazil.
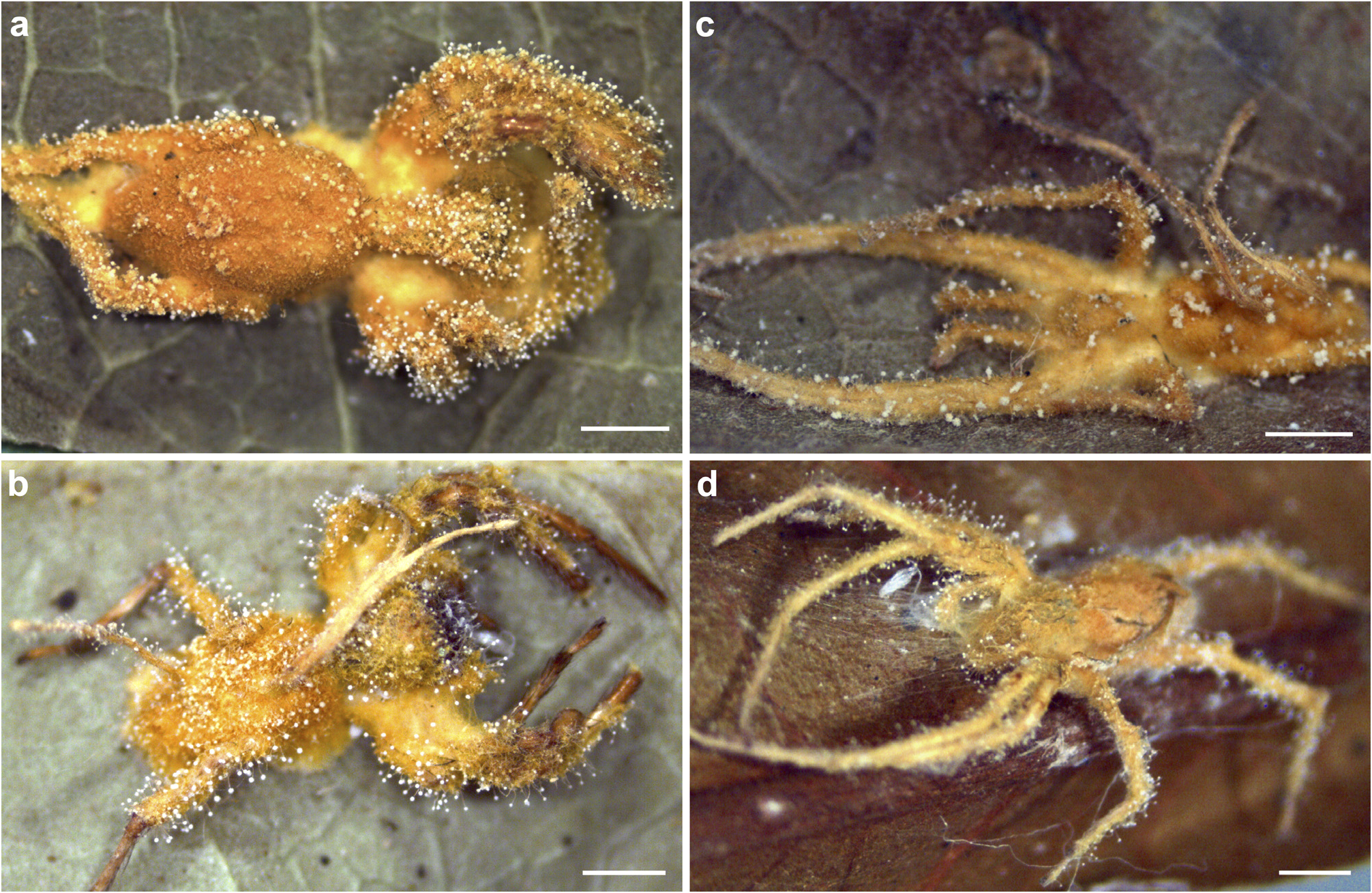
Hosts: —Spider species of the families Corinnidae (6.5% of the collected hosts)( Fig. 2a, b View FIGURE 2 ) and Anyphaenidae (87% of the collected hosts), including M. pacoti (comprising 69% of the hosts)( Fig. 2c, d View FIGURE 2 ); other 6.5% of the hosts were not identified. Host size 1.51–5.24 mm (min–max) long (N=14). Hosts were found attached to the abaxial surface of leaves in plants up to approximately 3 m high above the soil. No specific fungal structure attaching the host to the substrate was found.
Description: — Mycelium golden-yellow, completely covering the spider host ( Fig. 1a, k, l View FIGURE 1 ). Synnemata goldenyellow, solitary or up to three, 0.81–4.92 mm long × 0.09–0.27 mm wide (N=16), with a short stipe and fertile area ( Fig. 1a View FIGURE 1 , arrow), which consists of a network of anastomosed hyphae, hyaline, septate, terminating in an inflated fertile tip ( Fig. 1b, h View FIGURE 1 ). Conidiophores numerous, white, arising directly from the subiculum or produced along the synnemata, scattered, verrucose, hyaline, 86.56–319.88 (434.97) µm long × 6.52–17.64 µm wide at the base (N=33), aspergillate, narrowing to a slender apex (3.12–8.08 µm wide), and terminating in a swollen vesicle ( Fig. 1c View FIGURE 1 ). Conidial heads globose, white, 28.84–47.94 µm diam. (N=46). Vesicles ellipsoidal, (6.63) 8.53–15.57 µm long × 5.95–10.76 µm wide (N=15), hyaline, smooth, bearing several metulae ( Fig. 1d View FIGURE 1 ). Metulae ellipsoidal to obovoidal, hyaline, numerous, 5.47–12.27 µm long × 3.56–10.08 µm wide (N=130), borne on vesicle. Phialides cylindrical, 5.10–8.95 µm long × 1.5–3.57 µm wide (N=140), often apically thickened, smooth-walled, hyaline ( Fig. 1d View FIGURE 1 ). Conidia ellipsoidal, according to the length/width ratio ( Largent et al. 1977), with apiculate ends, smooth-walled, hyaline, 2.54–3.97 µm long × 1.78–2.90 µm wide (N=51) ( Fig. 1e, j View FIGURE 1 ). Granulomanus -like synanamorph conidiogenous cells (arrow) at the bases of conidiophores ( Fig. 1g View FIGURE 1 ). Conidiophores observed at the surface of the host body, especially on the legs ( Fig. 1f, i View FIGURE 1 ). Conidial head hemispherical, bearing ellipsoidal vesicle and metulae, distinct from the spheric conidial head found in most conidiophores. Conidia cylindrical to filiform ( Fig. 1f View FIGURE 1 ), 8.67–18.82 µm long × 0.79–1.56 µm wide (N=23). Sexual morph not observed.
Additional specimens examined: — BRAZIL. Ceará: Pacoti, Sítio São Luís, Trilha do Purgatório, 910 m elev., 4°13’21” S, 38°53’35” W, 21 December 2019, on spider M. pacoti, Alves, J. E. R. (#01; 04; 05; 06; 09; 10; 11; 12; 15; 16; 21; 22; 23; 27; 29; 30; 31; 33; 34; 35; 41; 42). Ibid. on spider Corinnidae, Alves, J. E. R. (#24; 25; 40; 43). Ibid. 18 January 2020, on spider M. pacoti, Alves, J. E. R. (#124; 125; 126; 128; 130). Ibid. 30 June 2021, on spider M. pacoti, Fonseca, E. O. (#08; 09, 10, 12). Paratype: Ibid. 03 April 2022, on spider M. pacoti, Fonseca, E. O. , deposited at the collection Herbário da UNILAB, (HU), HU1275, Collector number: EOF001.
Specimens sequenced directly from the host: LBMCF0003: spiders #01, 25, 33, 125, 128; LBMCF0004: #34, 35, 124, 126; LBMCF0006: #04; 06, 11, 15, 23, 31; LBMCF0007: #05, 22, 36, 29, 130; LBMCF0008: #04, 27; LBMCF0009: #30; LBMCF0010: #41.
Molecular phylogeny
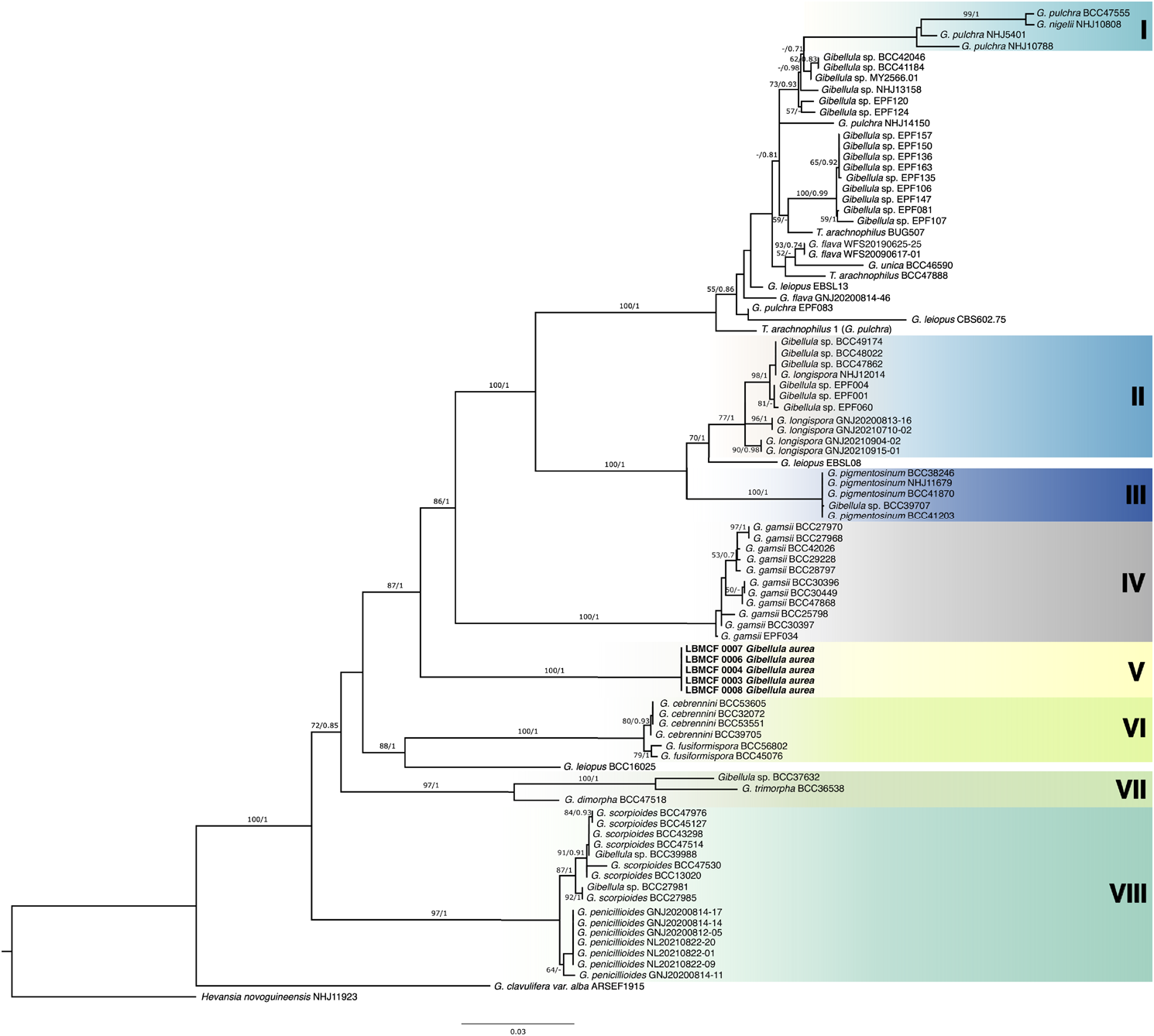
We obtained sequences of the genomic regions SSU (n=3), LSU (n=3), TEF (n=7), and RPB2 (n=4) from specimens of G. aurea using DNA directly extracted from synnemata ( Table 3 View TABLE 3 ). The 91 sequences of Gibellula from this study and previous studies included in the analyses clustered in eight clades, most exhibiting high support for both phylogenetic searches ( Fig. 3 View FIGURE 3 ; Fig. S1 View FIGURE 1 ). The phylogenetic trees generated by ML ( Fig. 3 View FIGURE 3 ) and BI ( Fig. S1 View FIGURE 1 ) using the combined dataset showed similar topologies for most of the clades, with posterior probabilities and bootstrap values higher than the threshold set (ML>50%, BI>0.70); especially for those specimens identified at the species level. The newly proposed species G. aurea clustered in a clade with different support for each topology, which displayed a bootstrap value of 87% and posterior probability of 1 (Clade V; Fig. 3 View FIGURE 3 ). Furthermore, nine species formerly described clustered in delimited clades ( Fig. 3 View FIGURE 3 ): G. longispora (Clade II), G. pigmentosinum (Clade III), G. gamsii (Clade IV), G. cebrennini / G. fusiformispora (Clade VI), G. dimorpha / G. trimorpha (Clade VII), and G. scorpioides / G. penicillioides (Clade VIII).
Intra- and interspecific genetic variation
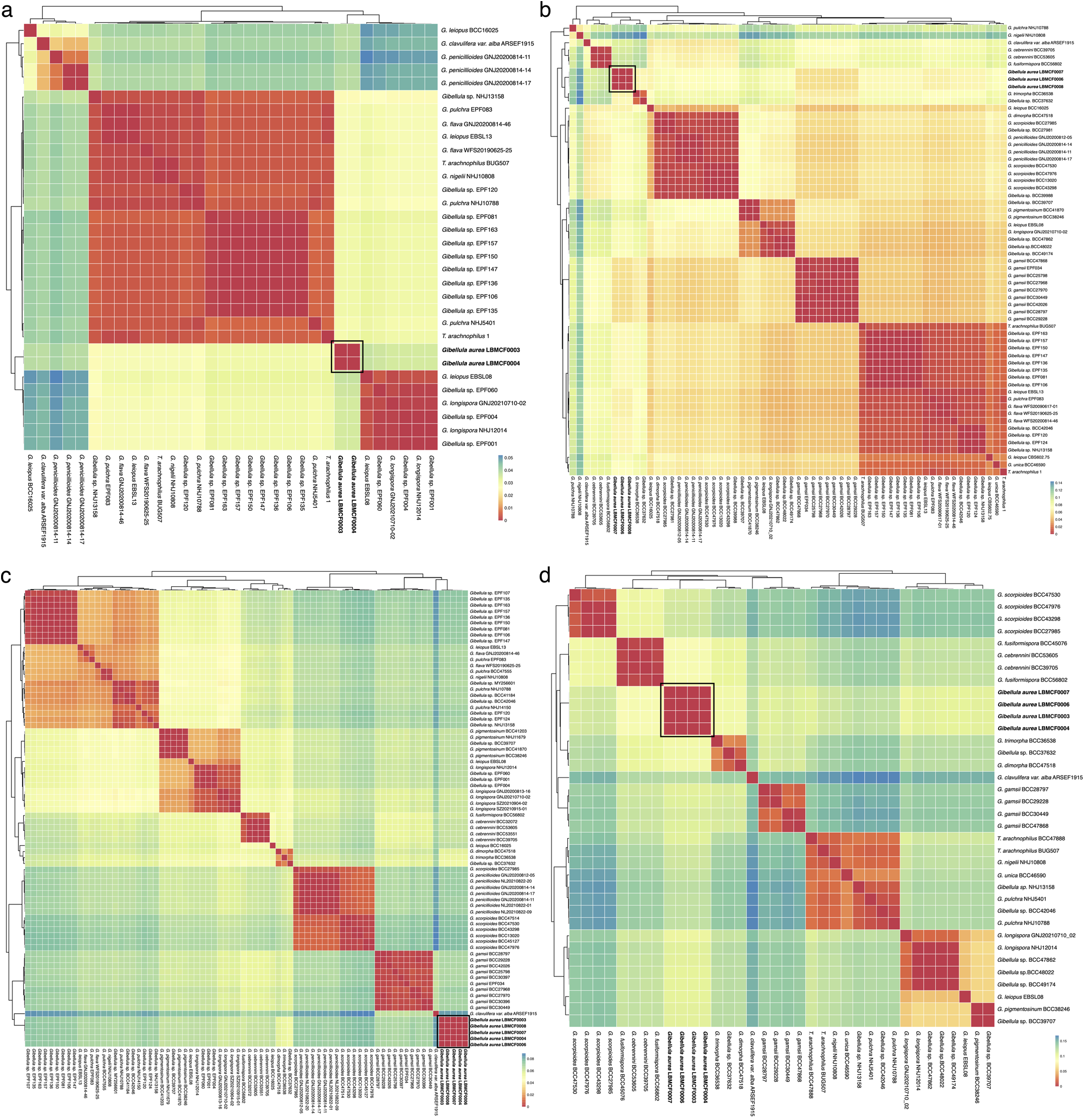
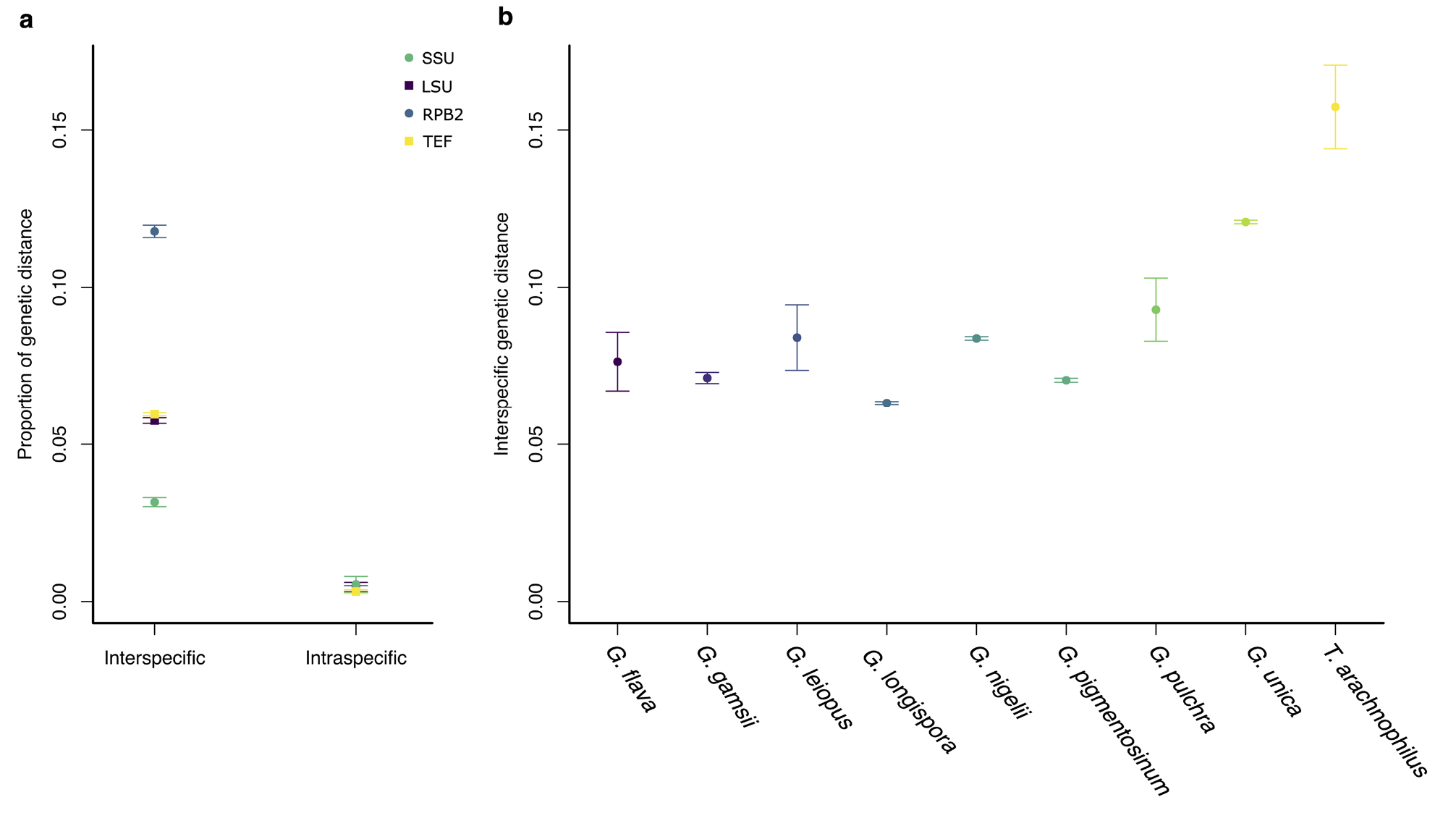
The specimens of G. aurea displayed lower intraspecific than interspecific distances for the four genomic regions (SSU: F (1,118) =81.69, p<0.001; LSU: F (1,593) =961.984, p<0.001; TEF: F (1,1324) =5632.558, p<0.001; RPB2: F (1,210) = 2559.837, p<0.001; Fig. 4 View FIGURE 4 ), which confirms the hypothesis of a new species initially suggested by morphological and phylogenetic analyses. Species that have been recently described with sequences for most of the genomic regions ( Chen et al. 2021, 2022; Kuephadungphan et al. 2019, 2020, 2022) also showed lower intraspecific than interspecific distances for the genomic markers shown here. Some sequences described only at the genus level were closer to described species, such as G. pulchra and G. leiopus , which might help to clarify the identity of these specimens ( Fig. 4 View FIGURE 4 ). The dataset of the RPB2-coding gene displayed higher genetic distances among the markers for the genus ( Fig. 5 View FIGURE 5 ; Table S3 View TABLE 3 ) but still displayed a barcode gap. Moreover, the LSU and TEF regions showed better-delimited groups.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |