
Metabelba (Pateribelba) sphagni Strenzke, 1950
|
publication ID |
https://doi.org/ 10.5281/zenodo.279150 |
|
DOI |
https://doi.org/10.5281/zenodo.6194472 |
|
persistent identifier |
https://treatment.plazi.org/id/03DF87DA-D15A-FF8F-11A5-F8C148ACFE80 |
|
treatment provided by |
Plazi |
|
scientific name |
Metabelba (Pateribelba) sphagni Strenzke, 1950 |
| status |
|
Metabelba (Pateribelba) sphagni Strenzke, 1950 View in CoL
Synonymy: Metabelba sphagni: Strenzke, 1950 ; Miko (2006). Metabelba lanceolata van der Hammen, 1952 (holotype documented in our Fig. 23 View FIGURE 23 ).
Diagnosis. Two pairs of centrodorsal tubercles (Da, Dp) present, well developed, Dp hidden under anterior margin of notogaster in dorsal view; postbothridial (Ba, Bp) and lateral (La) tubercles absent. Parastigmatic apophyses (Sa, Sp) of equal length, conical, pointed with convergent tips. Sensillus (ss) long with flagellate tip; covered with minute barbs in distal 1/2, flagellate tip virtually smooth. Interlamellar seta (in) almost as long as sensillus, thin with flagellate tip, covered with minute barbs in distal part. Notogastral setae inserted on minute protuberances, c1-c2, la -lp and h1-h3 strong, erect, spiniform, directed in radial pattern, smooth, darkly pigmented with hyaline base; c2- h2 shorter than mutual distance between the insertions of the two respective setae in pair. Discidium well developed, spiniform, straight, directed laterad. Cerotegument with long filamentous excrescences. Adults often carrying gastronotic exuviae (scalps).
Type material examined. Collection of Karl Strenzke, Senckenberg Naturmuseum, Frankfurt am Main, Germany: holotype ( SMF 14439) and 2 paratypes ( SMF 14441), individually mounted in 3 permanent mounts labelled as " Metabelba sphagni, Holstein 1949 , Nr 902". All three individuals are females mounted dorsal side upwards with legs stretched, and are cleared and undamaged, allowing detailed study. One of the paratypes has gastronotic exuviae on notogaster. The ventral side could not be studied in detail because of the relatively thick slide.
Collection of Leendert van der Hammen, Naturalis National Museum of Natural History Leiden, the Netherlands: permanent slide with 1 specimen labelled as “ Metabella lanceolata v.d. Hammen Holotype. l ad. Faure., Lattrop Bergvennen 22-IV-1949 Sphagnum monster A09.” Habitat data from van der Hammen (1952): A19. Sphagnum cuspidatum Ehr. and Sphagnum magellanicum Brid. from the border of the pool.The specimen (male?) is crushed, strongly damaged and cleared, the tarsi and tibiae of all legs and many of notogastral setae are missing.
Further material examined. Collection of Leendert van der Hammen, National Museum of Natural History Leiden, the Netherlands: permanent slide with 1 crushed specimen (female) with remains of gastronotic exuviae on notogaster, labelled as “ Metabella cf. sphagni Strenzke l Ad. Faure., Markelo (Twente) 29-X-1948 Vochtige heide leg. L. v.d. Hammen monster A16”.
Collection of Miroslav Kunst, Dept. of Zoology, Charles University Prague, Faculty of Science, Czech Republic: 1 specimen (female) mounted in permanent slide, 5 specimens unmounted, preserved in alcohol, labelled as " M. lanceolata, M. Tisý , 27. 8. 58 ". Details of the locality and habitat from Kunst (1959): moist Sphagnum meadow. Kunst (1959) originally identified specimens from this locality as M. sphagni . Kunst (1968) reindentified them as M. lanceolata . We confirmed the identity with M. sphagni through comparison with the type series.
Private collection of Gerd Weigmann, Berlin, Germany: 1 female and 1 male, Hochmoor "Hechtdiebel" 2.5 km north of Glambeck, near Eberswalde, Brandenburg, Germany, 53° 1.846' N, 13°48.740' E, floating Sphagnum , May 1991, leg. Christine Kehl.
Freshly collected material: Czech Republic, south Bohemia, Třeboňsko Protected Landscape Area, peat bog Ruda, in the vicinity of the field station of Dept. of Zoology, Charles University Prague, about 3 km south of Veselí nad Lužnicí 49°9' 4" N, 14°41'34" E, about 420 m a. s. l., wet meadow with Sphagnum : 40 specimens, 12. 4. 2009, 1 specimen 12. 7. 2009; 13 specimens, 11. 9. 2009; 35 specimens, 27. 3. 2010, all leg. Jitka Vilímová. All specimens unmounted, preserved in 80% ethanol or pure glycerine. From this material voucher specimens will be deposited the following museum collections: 5 specimens in Soil Zoology Collection of Hungarian Natural History Museum; 5 specimens in Acarological Collections of Senckenberg Museum für Naturkunde Görlitz ( Germany), 5 specimens in National Museum of Natural History Naturalis, Leiden ( the Netherlands), 5 specimens in Arachnological Collections in Zoological department of National Museum in Prague ( Czech Republic), 49 specimens in the collection of J. Mourek, Dept. of Zoology, Faculty of Science, Charles University Prague ( Czech Republic); 5 specimens in private collection of L. Miko; 5 specimens in collection of F. Bernini, Department of Evolutionary Biology, University of Siena ( Italy); 5 specimens in private collection of Gerd Weigmann, Berlin ( Germany).
Redescription (adult). Figs. 17–22 View FIGURE 17 View FIGURE 18 View FIGURE 19 View FIGURE 20 View FIGURE 21 View FIGURE 22 .
Dimensions. Holotype (female): ventral length 448 μm, maximum notogastral width 296 μm; paratype (female): ventral length 448 μm, total notogastral width 300 μm. The mounted type specimens measured in dorsal view. Specimens from South Bohemia: ventral length 416–448 (434) μm, maximum notogastral width 280–304 (293) μm; n= 5. Detailed measurements of legs in Table 1.
Integument. Body colour of fresh specimens usually medium brown, mounted holotype and paratypes pale yellowish (probably cleared for study by Strenzke or vanished in the medium). Cerotegument with "wool-like" filamentous excrescences; excrescences dense and comparatively short on legs ( Fig 22 View FIGURE 22 B), anogenital area and anterior part of coxisternum, long and loose on prodorsum, sejugal area and around notogastral setae. Underlying cuticle mostly smooth, but faintly granular on apophyses. Gastronotic exuviae (scalps) often present on notogaster of adult. Muscle insertions (sigillae) distinct in holotype and freshly collected fully sclerotized specimens, but hardly visible in paratypes.
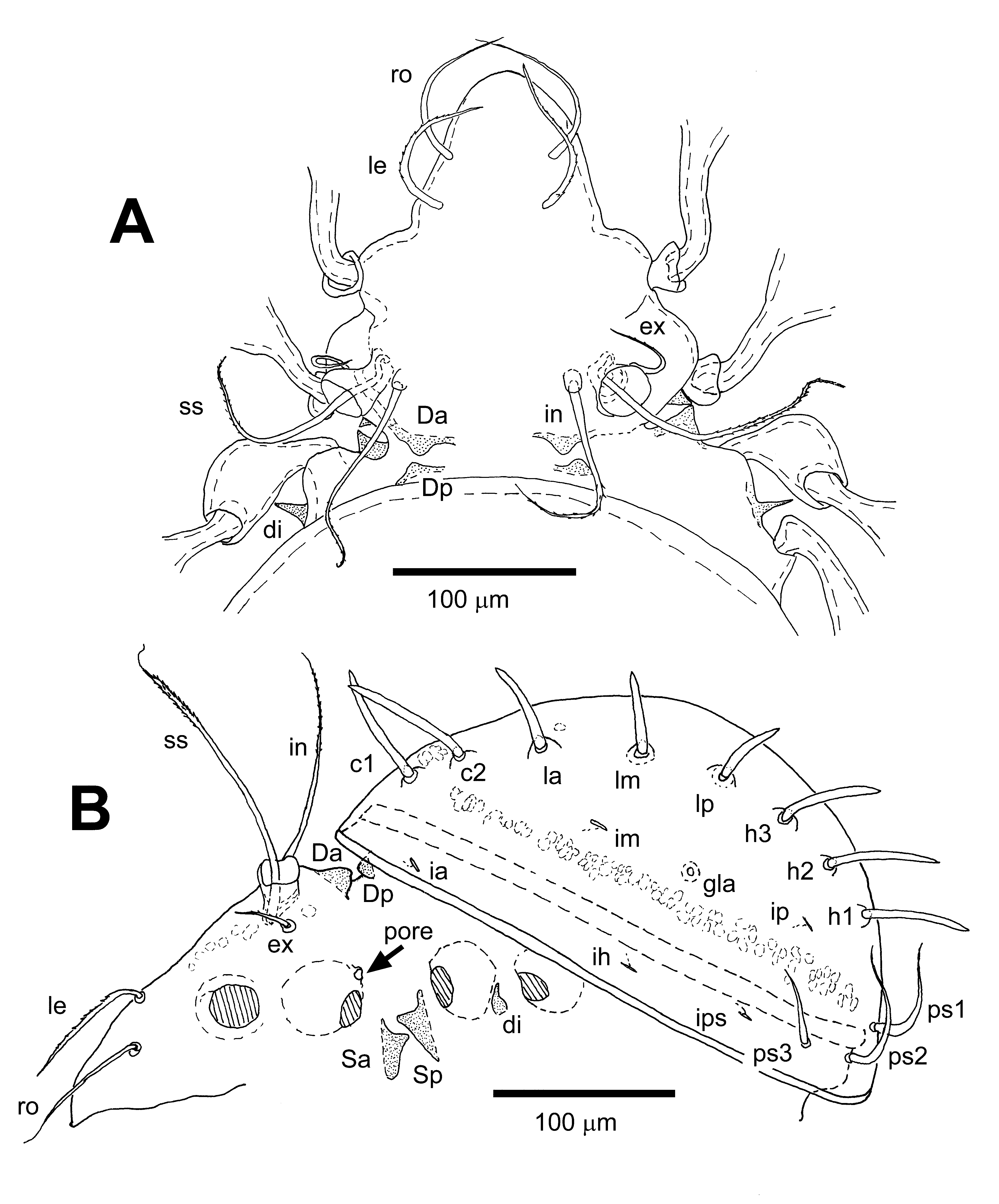
Prodorsum. Fig. 17 View FIGURE 17 , 18 View FIGURE 18 A. Prodorsum of roughly triangular shape, maximum width behind bothridia, about 0.7 of maximum notogastral width. Rostrum blunt with weakly defined nose-like protuberance on dorsal side, separated from rest of prodorsum by a paired shallow lateral indentation. Prodorsum broadened in dorsal aspect by a pair of distinct swellings at level of acetabula I. Apophysis P absent. Two pairs of centrodorsal tubercles (Da, Dp) present, well developed, Dp hidden under anterior margin of notogaster in dorsal view; postbothridial (Ba, Bp) and lateral (La) tubercles absent. Parastigmatic apophyses (Sa, Sp) of equal length, conical, pointed with convergent tips, only partly visible in dorsal view. Rostral (ro) and lamellar (le) setae comparatively long and strong, of approximately equal length. Seta ro slightly thinner than le, virtually smooth; inserted distinctly ventrolaterad from insertion of le, insertion of ro not visible in dorsal view. Seta le with a few minute barbs on outer curvature. Interlamellar seta (in) arising on minute apophysis, almost as long as sensillus, thin with flagellate tip, covered with minute barbs in distal half, flagellate tip virtually smooth. Exobothridial seta (ex) comparatively long, thin, curved anteromediad, covered with a few indistinct minute barbs on outer curvature. Sensillus (ss) long with short flagellate tip, sparsely covered with minute barbs in distal part. Bothridial funnel with slightly irregular rim, more developed on posterolateral side, normal for family.
Pair of distinct elongated fields of concentrated sigillae (muscle insertions) present between acetabula I and reaching anterior margin of bothridia; unpaired medial field of sigillae located between bothridia; pair of small sigillar fields located at anterior base of Da.
Notogaster. Figs. 17 View FIGURE 17 , 18 View FIGURE 18 B. Notogaster of broadly ovoid outline in dorsal view, hemispherical in lateral view. Broadest part of notogaster positioned slightly behind half of notogaster, anterior margin of notogaster covering dorsosejugal area in dorsal view. Notogastral setae of series c1-c2, la -lp and h1-h3 strong, erect, spiniform, directed in radial pattern; smooth, darkly pigmented with hyaline base; setae c2-h2 shorter than mutual distances in respective setal pairs. Mutual distance between insertions of c1 smaller than half mutual distance of insertions of c2. Setae ps1-ps3 comparatively long, thin, virtually smooth with attenuate tips, directed dorsolaterad, ps1 ≅ ps2> ps3. Circumgastric row of muscle sigillae (insertions of dorsoventral muscles) distinct, present laterally from setae c2- h3; groups of distinct sigillae present between insertions of setae c2 and posteromedially from seta h1. Pair of minute light spots present slightly posteromedial to insertions of setae la; smaller and less distinct pair of spots present medial from insertions of h3. Normal set of five lyrifissures (ia, im, ih, ip, ips) and opening of opisthonotal gland (gla) present on lateral part of notogaster. Notogaster often with gastronotic exuviae (scalps) arranged in pyramidal shape, but never with a compact mass of dirt. Cornicle k of nymphal gastronotic exuviae comparatively long, more or less straight ( Fig. 22 View FIGURE 22 D).
Gnathosoma . Fig.19 View FIGURE 19 . Subcapitulum normal for family: diarthric, with three pairs of thin and comparatively long setae (m> h> a) and two pairs of setiform processes (or1,2). Setae m, h, a covered with few minute barbs on outer curvature. Chelicera in shape and relative size typical of family, cheliceral seta cha with row of short barbs on outer (dorsal) curvature, seta chb with short barbs in distal third. Setation of chelicera not discernable, setal formula of pedipalp (from trochanter to tarsus, solenidion in parenthesis): 0-2-1-3-9(1).
Epimeral region. Fig. 19 View FIGURE 19 . Medial pit cp on coxisternum I not developed. Anterior tectum of podocephalic fossa extending laterally slightly under trochanter I, suture of podocephalic fossa almost perpendicular to body axis, without a tooth-like projection. Propodoventral (E2a, E2p) and ventrosejugal (Va, Vp) enantiophyses absent. Ventrosejugal furrow complete, body distinctly narrowed between acetabula II and III. Epimeral setal formula (I to IV): 3-1-3-4, normal for Metabelba . Muscle sigillae on epimeres distinct in most of studied specimens. Epimeral setae comparatively long, smooth, thin, with attenuate tips.
Anogenital region. Fig. 19 View FIGURE 19 . Discidium (di) well developed, spiniform, pointed, straight, directed laterad. Setal formula of anogenital region normal for the Damaeidae ( Grandjean 1960; Norton 1977b); genital g: 6; aggenital ag: 1; anal an: 2; adanal ad: 3; all setae comparatively long, thin with pointed attenuate tips. Lyrifissure iad oblique, divergent posteriad.
Legs. Figs. 20 View FIGURE 20 , 21 View FIGURE 21 AB. Legs monodactyl, moderately long, segments distinctly swollen in distal part. Leg IV slightly longer than body, leg I and III as long as or slightly shorter than body, leg II distinctly shorter than body (detailed measurements in Table 1). Leg setae normal for family, comparatively long and thin, pointed; most covered with minute barbs on outer curvatures. Dorsal seta (d) on femora strong, roughened with distinct barbs on outer curvatures. Leg setal formula identical with those of Metabelba pulverosa Strenzke, 1953 (see Norton 1977a) and M. denscanis n. sp. (see above). Solenidia ω 1,2 on tarsus I moderately long, tactile; φ 1 on tibia I and φ on tibia IV very long, tactile; ω 1,2 on tarsus II moderately long, setiform; φ 1 on tibia I about 3.5x longer than φ 2; φ on tibia IV slightly less than 2x longer than tibia itself. Solenidion φ slightly shorter than respective setae d on tibia II and slightly longer than respective setae d on tibia III. Solenidion σ as long as respective setae d on genu I and II and slightly shorter than respective seta d on genu III.
Geographical distribution and ecology. Metabelba sphagni is hitherto known from northern Germany: Schleswig-Holstein ( Strenzke 1950; Strenzke 1952) and Brandenburg ( Kehl, 1997); the Netherlands ( Siepel et al. 2009; unpublished record by van der Hammen — see our Material examined); Czech Republic: Central Bohemia ( Kunst 1959, 1962; Starý 2000a); Austria: Tyrol ( Jahn 1967; Schatz 1983); Switzerland: Swiss Jura Mts. ( Borcard, 1992); Sweden ( Tarras-Wahlberg 1961; Lundquist 1987) and Russia: Kola Peninsula (Krivoluckij et al., 1995).
The species seems to be comparatively rare and has been reported almost exclusively from Sphagnetum —peat bogs or moist meadows with predominating Sphagnum mosses—in relatively low altitudes above sea level ( Strenzke 1950; Kunst 1959, 1962; Tarras-Wahlberg 1961; Lundquist 1987; Kehl 1997). The recent findings from Ruda in South Bohemia and other specimens examined from museum collections (see Material examined) are from the same type of habitat. Similarily, M. lanceolata , junior synonym of M. sphagni was collected in wet Sphagnum on the border of a pool in the Netherlands (van der Hammen 1952). In contrast, Jahn (1967) reported M. sphagni from a larch forest ( Larix decidua ) in the altitude about 1100 m a.s.l. and Borcard (1992) reported it from a peat-bog in the altitude 1090 m a. s. l.
Most authors found only a few individuals of M. sphagni and the abundant population found in the peat bog Ruda in South Bohemia (see our Material examined) seems to be an exception. It is possible that the rarity of records of M. sphagni is caused by the fact that this type of habitat has been poorly studied.
Remarks. Metabelba sphagni is distinguished from other species of the subgenus mainly by the combination of two characters—presence of two pairs of well-developed centrodorsal tubercles (Da, Dp) and strong, almost straight and smooth spiniform notogastral setae, shorter than their mutual distance, which are inserted on distinct protuberances. In M. glabriseta , known from Ethiopia, two pairs of centrodorsal tubercles are also present, but the species has relatively longer notogastral setae. Moreover, it is considerably bigger in size and posterior ventrosejugal tubercle (Vp) is present. M. platynotus is another species with two pairs of centrodorsal tubercles, but it has very short notogastral setae and conical notogaster with flattened anterior part.
The possibility that Metabelba lanceolata , van der Hammen, 1952, may be a junior synonym of M. sphagni was suggested by Miko (2006). Subías (2008, 2009) stated the synonymy of both species as a fact, without providing a reference, probably influenced by Miko (2006). In this study we compared type specimens of both species and substantiated the synonymy, based on the following analysis:
1. The original description of M. lanceolata is brief and incomplete, based on a single crushed and strongly damaged individual (van der Hammen, 1952; see also our Material examined and Fig. 23 View FIGURE 23 ): tarsi and tibiae of all legs were missing, so that the presence of setae d on tibiae II and III could not be studied, therefore the author noted that the generic position of the species was uncertain. Moreover, many of notogastral setae were missing. Van der Hammen (1952) illustrated the specimen with strongly obovate notogaster strongly narrowing posteriad, which is very unusual in Damaeidae . We conclude that this was an incorrect and misleading reconstruction of the shape of the crushed specimen in permanent slide.
2. The main difference of M. lanceolata from M. sphagni should be the broadened—“lanceolate”- shape of notogastral setae (van der Hammen, 1952) and this character was adopted in diagnostic keys by subsequent authors (van der Hammen & Strenzke, 1953; Kunst 1961; Bulanova-Zachvatkina 1975). However, from the careful examination of the holotype of M. lanceolata , we conclude that the “lanceolate”- shape of notogastral setae is only an artefact, caused probably by intensive maceration of the specimen and subsequent strong compression of the setae by the cover glass in permanent slide. From our Fig 23 View FIGURE 23 B is evident, that in the seemingly "lanceolate" setae lm, lp, h3 and h2 the surface layer is detached from the core of the seta and flattened by the compression. In contrast, seta h1 has normal appearance as in M. sphagni .
The thickness of notogastral setae in M. sphagni is slightly variable, some specimens from South Bohemia having somewhat thicker setae than the type series ( Fig. 21 View FIGURE 21 CD). However, the “impression of thickness” of the setae depends also on the position of a given specimen during the observation — the setae appear to be thicker in dorsolateral view than in exactly dorsal position — as we demonstrate on Fig. 21 View FIGURE 21 CD.
3. Kunst (1968) found also the length ratio between trochanter and femur IV to be a distinguishing character between M. sphagni and M. lanceolata , the former species having trochanter IV shorter than femur IV and the contrary in the latter species. His assumption was not based on the study of type or topotypical material of either species, but on comparison of specimens from South Bohemia identified as M. lanceolata with original description by Strenzke (1950). However, we could not confirm this difference; all available specimens from the type series of M. sphagni had trochanter IV slightly longer than femur IV, in the holotype of M. lanceolata both segments were equally long and the ratio slightly varied among the material of M. sphagni from South Bohemia (see Table 1). The two species therefore cannot be distinguished according to the length ratio of trochanter IV and femur IV. It is possible, that Strenzke (1950) measured the given leg segments in a different way. The synonymy is supported by the fact that both species have been found in wet habitats, particularly peat bogs in lowland ( Strenzke 1950, van der Hammen 1952, Kunst 1968).






| SMF |
Forschungsinstitut und Natur-Museum Senckenberg |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |