
Homalocantha Mörch, 1852
|
publication ID |
https://doi.org/10.5252/z2011n4a7 |
|
persistent identifier |
https://treatment.plazi.org/id/03E05218-FF9A-FFC4-30EE-FC7DFC1CE7AF |
|
treatment provided by |
Felipe |
|
scientific name |
Homalocantha Mörch, 1852 |
| status |
|
Genus Homalocantha Mörch, 1852 View in CoL
TYPE SPECIES. — Murex scorpio Linnaeus, 1758 by monotypy.
COMMENTS ON THE SUBFAMILIAL ASSIGMENT
The genus Homalocantha should be placed in incertae sedis in the subfamilies Muricopsinae and Muricinae (see Barco et al. 2010). We place here Homalocantha in the Muricopsinae as did Vokes (1994), but this placement is provisional, waiting for new phylogenetic data based on molecular studies.
COMMENTS ON THE SPECIES OF THE GROUP
OF H. SCORPIO (LINNAEUS, 1758)
The two new species described here belong to a group of close species including H. scorpio (Linnaeus, 1758) , H. pisori d’Attilio & Kosuge, 1989 , H. vicdani d’Attilio & Kosuge, 1989 and H. dondani d’Attilio & Kosuge, 1989 . Historically, H. scorpio was defined on the basis of adult specimens, but the three other species were defined only on the basis of juvenile specimens
( d’Attilio & Kosuge 1989). Consequently, juveniles of H. scorpio were not represented or figured by d’Attilio & Kosuge (1989) and adults of the other three species have not been described. Since d’Attilio & Kosuge (1989), no worker was interested to document better the ontogeny of these species. The opportunity to study new material allows relating juveniles to adults and then to understand and define these species better from a growth series. Thus, it is the first time that a growth series is presented for these species ( Figs 1-6 View FIG View FIG View FIG View FIG View FIG View FIG ). Our observations are based on 225 specimens of H. scorpio , 32 specimens of H. pisori , 25 specimens of H. vicdani , 4 specimens of H. dondani , 5 specimens of H. granpoderi n. sp. and 29 specimens of H. ninae n. sp. Considering only adults morphologies, morphological differences could be interpreted as intraspecific variations, but young morphologies appear poorly variable and more differentiated than adults. For example, juveniles of H. scorpio (e.g., Fig. 3A View FIG ) display few variations regarding their cords, their shape and their colour and differ clearly from the juveniles of H. granpoderi n. sp. ( Fig. 1 View FIG ). So, our experience based on numerous specimens suggests that, for this group of Homalocantha , young morphologies characterize species better than the adult morphologies do. On the evolutionary point of view, ontogenic changes affecting younger morphologies than adult morphologies correspond to a heterochronic process called early innovations ( Dommergues et al. 1986; Videt & Néraudeau 2003). On the other hand, the Philippines hotspot is identified as one of the world’s biologically richest areas and it is not surprising to find several muricid species of the same genus in this area. In comparison to the Philippines archipelago, the São Tomé and Principe Islands (West Africa), a smaller area with a lower species richness, contains no more than six species of Muricopsis ( Rolán & Fernandes 1991; Houart 1996, 2005; Rolán & Gori 2007; Houart & Gori, 2008): M. rutilus mariangelae Rolán & Fernandes, 1991 , M. matildae Rolán & Fernandes, 1991 , M. principensis Rolán & Fernandes, 1991 , M. delemarrei Houart, 2005 , M. hernandezi Rolán & Gori, 2007 and M. ( s.s.) testorii Houart & Gori, 2008.
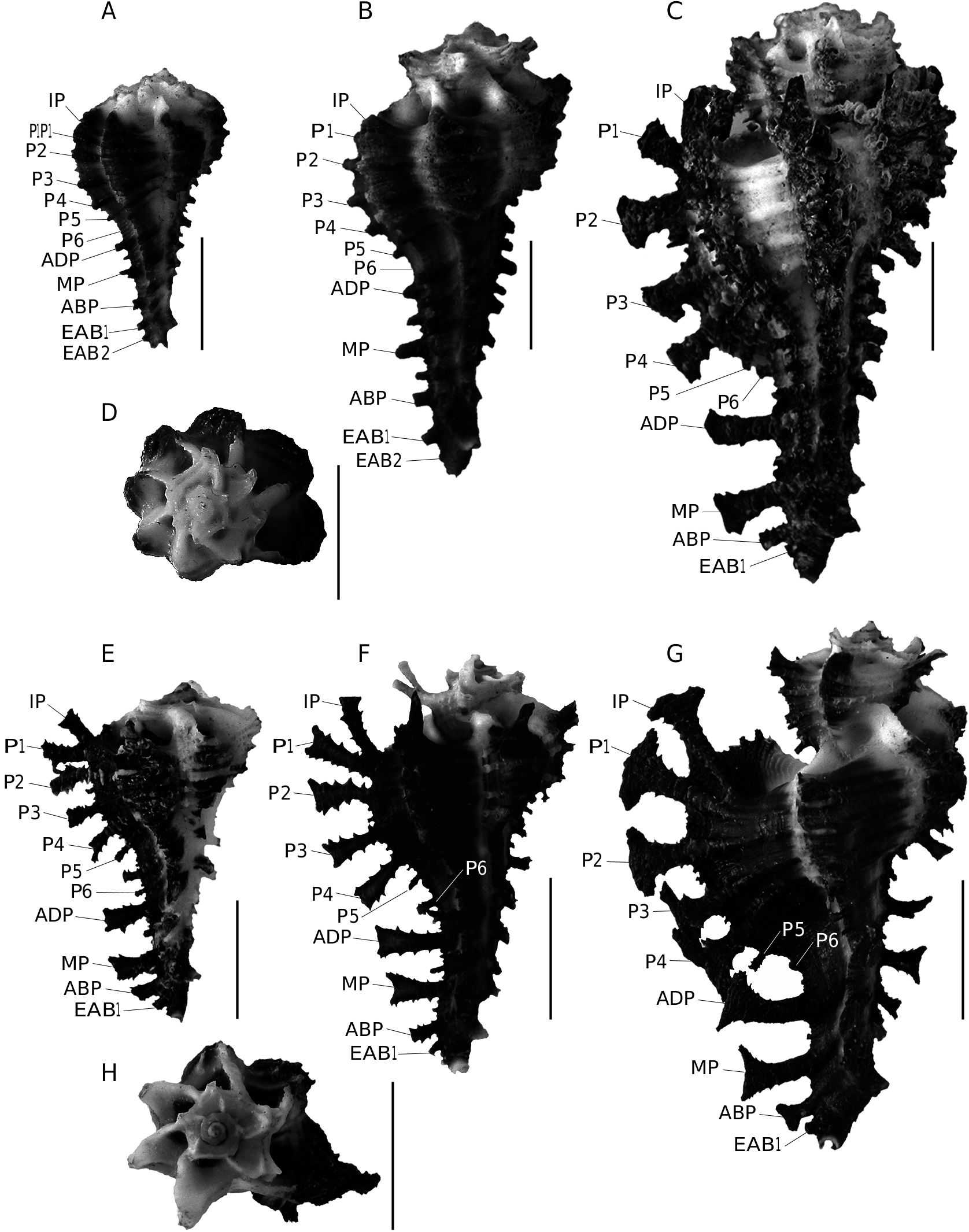
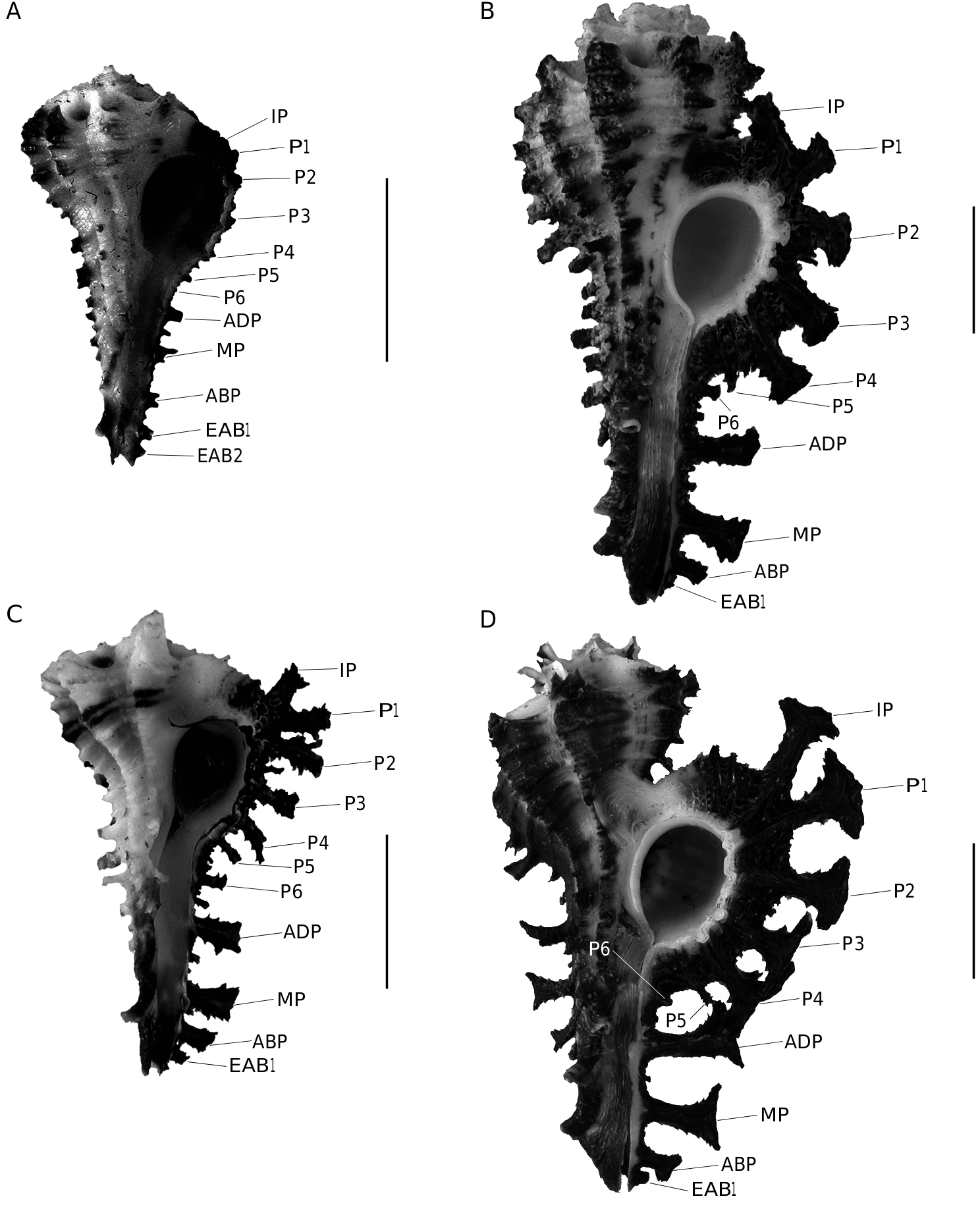

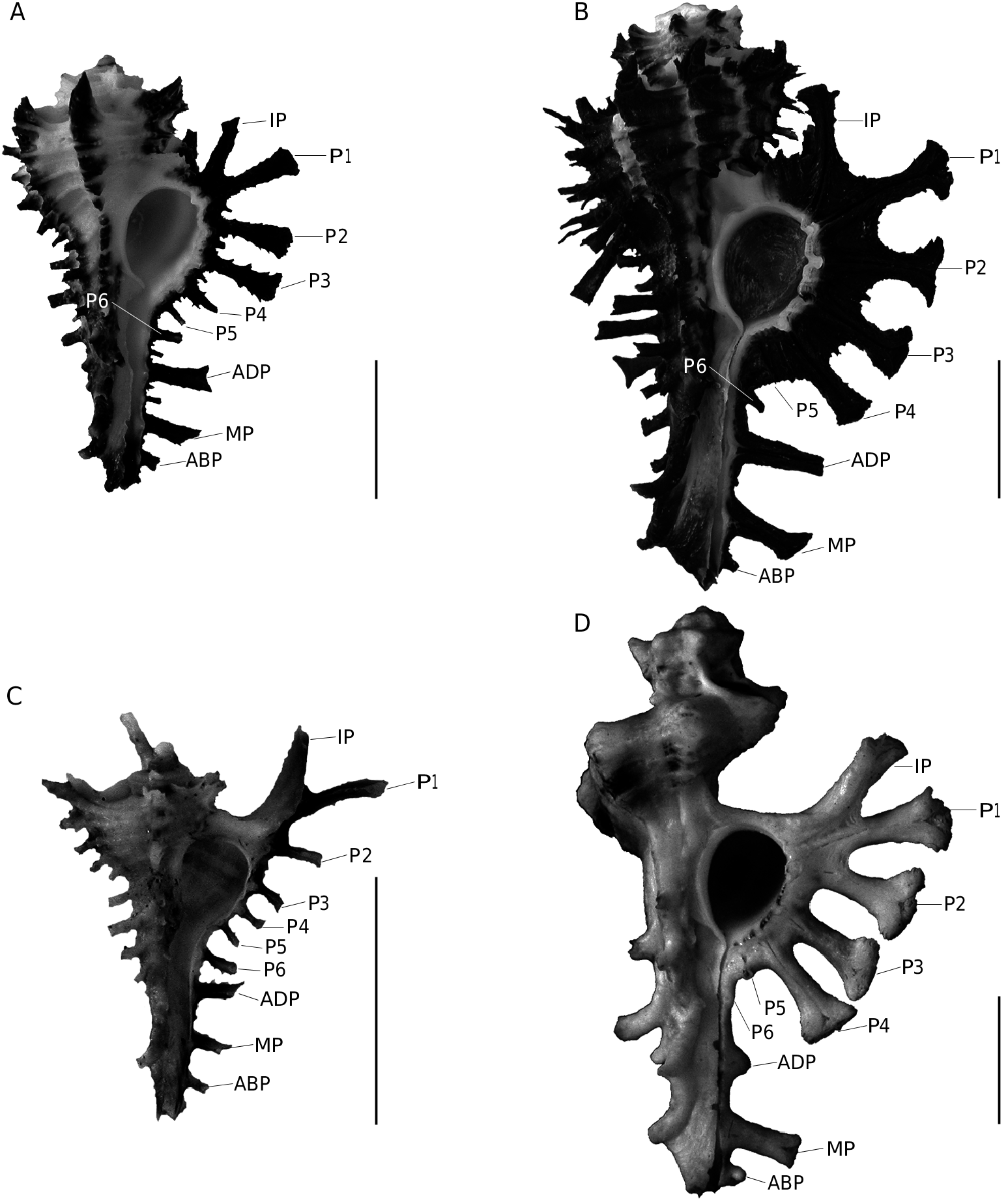


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.