
Parahelicops annamensis Bourret, 1934
|
publication ID |
https://doi.org/10.11646/zootaxa.3948.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:064A12F0-5742-4828-8D26-85AD7B226091 |
|
DOI |
https://doi.org/10.5281/zenodo.6096906 |
|
persistent identifier |
https://treatment.plazi.org/id/03E0879F-2472-FFA4-FAD0-6C63FB87F9B4 |
|
treatment provided by |
Plazi |
|
scientific name |
Parahelicops annamensis Bourret, 1934 |
| status |
|
Parahelicops annamensis Bourret, 1934
( Fig. 3 View FIGURE 3 A–D, 4A–C)
Parahelicops annamensis Bourret, 1934b: 170 , Fig. 2 View FIGURE 2 . Type locality. “Bana, station d’altitude située à 1500 m d’altitude au bord de la Baie de Tourane”, now Ba Na Nature Reserve, Da Nang City, Vietnam. Holotype. MNHN 1938.0117, adult male; collected by René Bourret, 1934.
Material examined ( 15 specimens). Vietnam. Ha Tinh Province. AMNH 147129, Po Mu forest ( 18°20'26"N, 105°14'13"E), Rao An, Huong Son District, elev. 870 m; ZISP NLO- 2478, Rao An, elev. 300 m. Quang Binh Province. ZFMK 86457, ZISP NLO- 2714, IEBR A.2013.16–17, Phong Nha - Ke Bang National Park, Hoa Son Commune, Minh Hoa District, 17°42.654'N, 105°52.708'E, elev. 527 m. Quang Tri Province. ZISP NLO- 3257, Ban Cup, Huong Hoa District, 16°57.434'N, 106°35.329'E elev. 490 m. Thua Thien-Hue Province. ZFMK 83513, Bach Ma National Park; ZISP NLO- 2790, Bach Ma National Park, elev. 1,350 m. Da Nang City. MNHN 1938.0117 ( holotype of Parahelicops annamensis ), “Bana”, now Ba Na Nature Reserve, elev. 1,500 m; ZISP NLO- 2831, Ba Na Nature Reserve, elev. 1,400 m. Kon Tum Province. IEBR 371, ZISP NLO- 3435, Mang Canh village (14°41'950N, 108°14'642E), Kon Plong District, elev. 1,200–1,250 m. Laos. Xékong Province. FMNH 258637, Xe Sap National Biodiversity Conservation Area, 16°04'10''N- 106°58'45"E, Kaleum District, elev. 1,280–1,500 m; NCSM 78632, Phou Ajol, 15.68516°N- 107°1981°E, Dakchung District, elev. 1,450 m.
Taxonomic comments. For a long time, this species has been known from only the holotype collected in central Vietnam. The second known specimen, and first recorded from Laos, was described by Stuart (2006). Subsequently, more specimens became available, especially from central Vietnam. This species is monotypic.
Diagnosis. A species of the genus Parahelicops characterized by the combination of (1) body elongate in males and females; (2) tail long with a ratio TaL / TL at least equal to 0.30, (3) maxillary teeth 28–34, the last two moderately enlarged; (4) DSR 15 or 17 at midbody, moderately keeled at midbody, strongly keeled posteriorly, usually smooth on the 1st dorsal scale row; (5) scales around the base of the tail very strongly keeled; (6) background colour chestnut brown, dark brown or dark greyish-brown; (7) dorsal pattern with isolate, bright orange or rusty-red blotches (cream or yellowish-ochre in preservative), usually horizontally elongate or vertically elongate as crossbars anteriorly, then becoming irregular, broken spots posteriorly; (8) postocular streak short, narrow, oblique, reaching the neck behind the corner of the mouth; (9) a broad, oblique and elongate orange or rusty blotch (cream or yellowish-ochre in preservative) present on the nape; (10) SL dark as the upper head surface, without defined spots or blotches, (11) venter pale in its central half with outer tips of ventral scales very dark; (12) eye moderate, 1.2–1.4 times the distance between the lower margins of eye and of lip; (13) VEN 158–172, SC 116– 146, paired; (14) internasals narrowed anteriorly; (15) SL 8 or 9; (16) prefrontal scale often single; and (17) anterior temporal single.
With its typical dorsal and ventral patterns and its long tail, Parahelicops annamensis cannot be confused with any species of the genera Opisthotropis , Paratapinophis , Amphiesma and Rhabdophis recorded from Vietnam and Laos. It is however much similar with Pararhabdophis chapaensis Bourret, 1934 (see below). It can be distinguished from this latter species by its pale venter, at least in its central area, vs. the venter entirely black in Pararhabdophis .
In collections, specimens of Parahelicops annamensis have been misidentified as Amphiesma modestum ( Günther, 1875) , an Indo-Himalayan species unknown east of Myanmar (our data, in prep.). Nevertheless, A. modestum has 19 DSR at midbody, 143–163 VEN, scales in the region of the base of the tail feebly keeled, and an ochre or pale brown venter.
Description. The body is moderately robust but elongate (more gracile in males) and cylindrical; head oval, rather short, moderately distinct from the rather thick neck, flattened anteriorly; snout long, 2.0–2.2 times as long as horizontal diameter of eye, flat, blunt or slightly rounded seen from above, blunt and subrectangular seen in profile, no canthus rostralis; nostril directed distinctly dorsolaterally, small, rounded, piercing in the middle of the divided nasal; eye moderately sized or small, diameter about 1.2–1.4 times the distance between the lower margins of eye and of lip, with a round pupil; tail long, thin and tapering.
The maximum known length is 796 mm for a male (SVL 528 mm, TaL 268 mm; specimen ZISP NLO-3257). The longest known female is 685 mm long (SVL 465 mm, TaL 220 mm; ZISP NLO-2831).
Ratio TaL / TL 0.297–0.337, without clear sexual dimorphism ( 6 males: 0.301–0.337; 3 females: 0.297– 0.321).
Maxillary teeth. Total of 28–34 maxillary teeth, gradually enlarged in a continuous series, the last 2 moderately enlarged, without diastema.
Body scalation. DSR 17–19 – 15–17 – 15–17; dorsal scales rhomboedric, normal in shape, distinctly keeled on 2nd–10th rows, moderately on the anterior part of the body, strongly posteriorly, very strongly in the region of the vent and on the base of the tail; scales smooth on 1st row or feebly keeled posteriorly; VEN 158–172 (plus 1 or 2 preventrals), with a sexual dimorphism (see below); SC 116–146, all paired, with a sexual dimorphism (see below); anal plate divided; ratio VEN / SC 1.18–1.43.
The number of DSR is quite variable in this species. In the 13 available specimens, the formulae are: 18–15–15 ( 1 specimen), 19–15–15 (3), 17–17–17 (1), 18–17–15 (2), 18–17–17 (2), and 19–17–17 (4).
Scale row reductions: first reduction (18 or 19→17) at VEN 56–89; second reduction (17→15) at VEN 98– 128.
Head scalation. Complement of upper head scales complete but with prefrontals variable, including 2 internasals, 1 or 2 prefrontals (see below), 2 supraoculars, 1 frontal, and 2 parietals. Rostral hexagonal, wider than high, barely visible from above; nasal subrectangular, elongate, vertically divided by a furrow, with the fore and hind parts of similar size, altogether about 1.3–1.5 times longer than high; internasals distinctly subtriangular, elongate, in broad contact with each other, 1.2–1.4 times longer than wide, distinctly narrowed anteriorly with anterior margin about 0.4–0.5 times the width of the posterior margin; prefrontal 1 (in 10 / 13 specimens), wide and shield-like, or 2 (3 / 13), large, subrectangular, broader than long, in both cases 0.9–1.2 times as long as internasals, in broad contact with loreal; supraocular 1 / 1, entire, subtriangular, rather broad, 1.8–2.2 times longer than wide, slightly narrower than internasals, about 0.5 times as wide as frontal; frontal hexagonal, shield-like, rather wide but short, its apex directed posteriorly, 1.0–1.2 times longer than wide, 1.7–2.0 times longer than prefrontals; parietals large, long and wide, 1.7–2.1 times longer than frontal and in contact for a length 1.2 times as great as the frontal length; loreal scale 1 / 1, large, rectangular, elongate, 1.5–1.8 times longer than high, in broad contact with the nasal; SL 8 or 9, 1st as long as high, 2nd–5th longer than high, other SL as long as high, 1st–2nd SL, small and short or, exceptionally 1st–3rd SL, in contact with nasal, 2nd–3rd or 3rd–4th SL in large contact with the loreal, 4th–5th SL, or exceptionally solely 4th, only 5th, 4th–6th or 5th–6th SL, entering orbit; 6th–7th or 7th–8th SL largest; preoculars 2 / 2 in all known specimens, subequal; postoculars 2 or rarely 3, small, the upper one largest; anterior temporal 1, narrow and elongate, in all known specimens with complete temporal formulae as 1+1+2 or rarely 1+2+2 or 1+3 (this latter met only in the type); IL 10 (rarely 8 or 9), 1st pair in contact behind the mental, 1st–5th IL in contact with anterior chin shields, 5th IL largest; mental triangular; anterior pair of chin shields equal to or slightly longer than posterior shields.
Coloration and pattern. In preservative, the dorsal and lateral surfaces of body are dark chestnut brown, dark brown or dark greyish-brown, darker above than on the sides, either nearly uniform or, more usually, profusely variegated with darker brown, diffuse blotches, larger on the upper parts of the sides than on the back; in 3 of 15 specimens, a faint dorsolateral stripe is present on 5th–7th dorsal rows, barely paler than the background color and poorly distinct; on each side, a series of about 50 to 70 distinct dorsolateral blotches, cream, ochre, yellowishbrown, pinkish-brown or orange-brown on 5th–7th dorsal rows, separated from each other by 2–3 dorsal scales, forming a discontinuous dorsolateral stripe; these blotches are large, thick and horizontally elongate on the fore part of the body from behind the neck to the first anterior quarter to third of the body, or, often, vertically elongate on a short part of the body up to forming a few vertical, more or less narrow crossbars reaching downwards to the level of the 3rd scale row; progressively these blotches are reduced to irregular or broken spots, or are absent posteriorly; a short, cream ventrolateral stripe on the anterior part of the body, up to 20th to 30th VEN. Upper surface of tail as the body, either uniform or with only a few pale dots.
Head dark chestnut-brown, more or less variegated above with ochre yellow or pale yellowish-brown, often with numerous minute scattered dark dots; an ochre yellow or pale yellowish-brown sagittal line present, but faint, or absent; last three posterior supralabials irregularly and faintly edged with ochre-yellow anteriorly; a short, narrow or relatively broad, oblique, more or less distinct postocular streak on anterior and lower posterior temporals, and on top of 8th and 9th SL, reaching the neck behind the corner of the mouth and continuing as a large, cream or yellowish-ochre oblique, elongate blotch on the nape, connecting the posterior end of the postocular streak with the first dorsolateral blotch. The throat, chin and IL are cream or pale yellowish-ochre, more or less heavily spotted with dark brown spots; IL heavily marbled or spotted with dark brown. Iris deep black in life.
Venter cream, pale yellowish-ochre or light brown, with outer tips of ventral scales dark brown or dark greyish-brown, progressively widening posteriorly; often some irregular dark spots scattered on the pale part. Under surface of tail as the venter but the pale central part narrower than the dark outer parts anteriorly, the tail becoming progressively entirely dark below.
It should be noted that the holotype has a venter much darker than other specimens (perhaps due to an inadequate preservation), but the bicolour ventral pattern typical of this species is nevertheless present.
In life, the background coloration is bright chestnut-brown or dark purplish-brown; the dorsolateral blotches are bright orange or rusty-brown, a colour often still seen in recently preserved specimens. The postocular streak is yellow turning to orange on the neck. Lastly, the pale area of the venter is orange or reddish-pink.
Hemipenis. In situ, it is short, reaching the level of the 8th SC, rather thin, and single although forked at its tips, the tips stemming from the face bearing the sulcus. The organ is calyculate on its lower proximal part, along about 1/6 of the organ length, then covered with medium-sized spines; some spines are larger and hooked near the limit between the calyculate and the spinose areas and on the most proximal part, shorter but more densely spaced towards the distal area. The sulcus is not conspicuous and mostly straight.
Sexual dimorphism. It is expressed in (1) the difference in the number of VEN: 7 males: 168–172 ( x = 169.7, s = 1.7); 6 females: 158–167 ( x = 164.2, s = 3.4); (2) difference in the number of SC: 6 males: 126–146 ( x = 138.7, s = 6.9); 3 females: 116–126 ( x = 119.7, s = 5.5); and (3) difference in the position (counted in SC) to the reduction from 6 to 4 scale rows around the tail of SC: males: 12th–16th SC; females: 4th–7th SC.
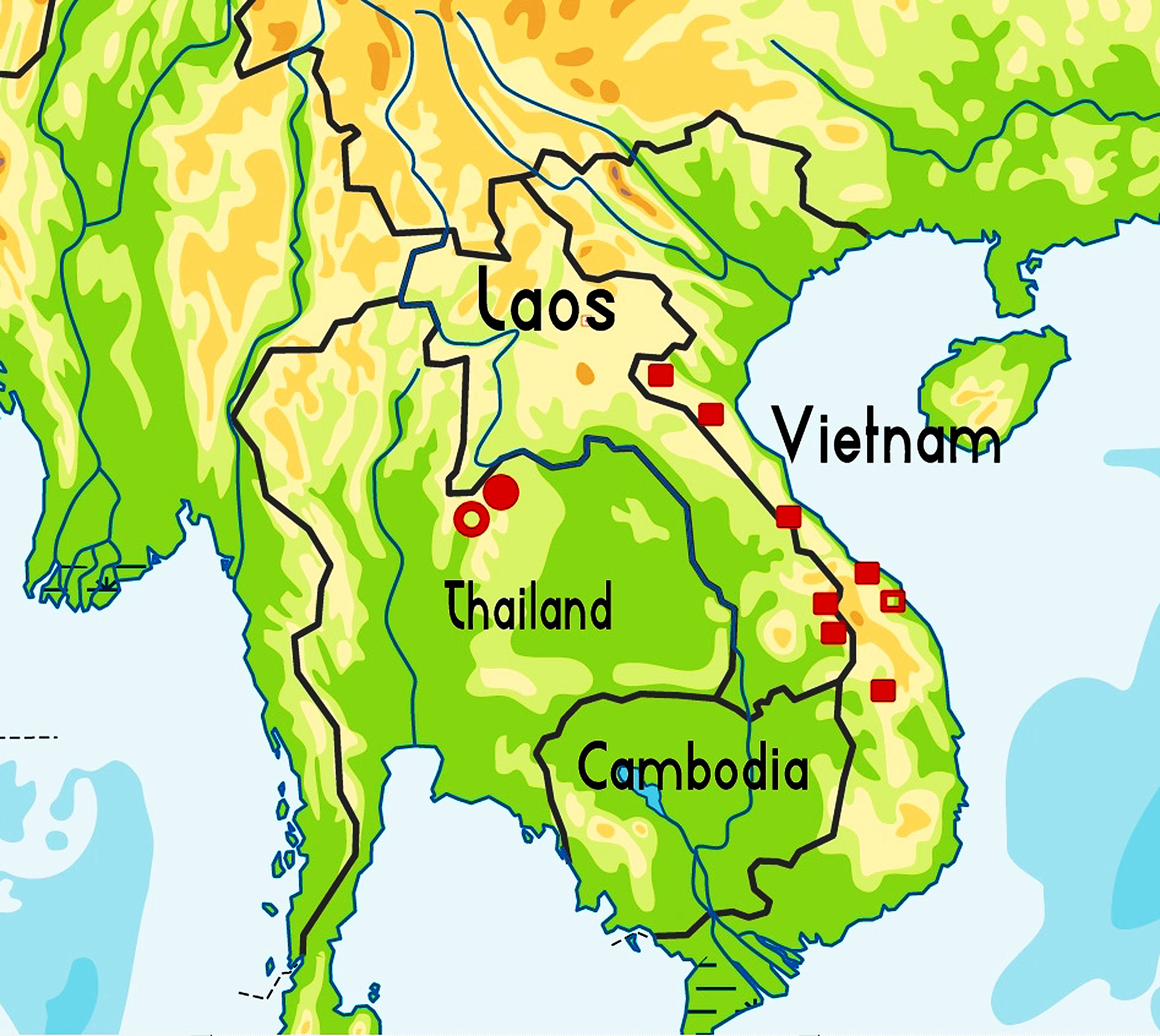
Distribution ( Fig. 2 View FIGURE 2 ). Vietnam. This species is known from the centre of the country, in the following provinces (from north to south): Ha Tinh (Huong Son District); Quang Binh (Phong Nha - Ke Bang National Park, Minh Hoa District); Quang Tri (Huong Hoa District); Thua Thien-Hue (Bach Ma National Park); Da Nang City (Ba Na Nature Reserve), and Kon Tum (Kon Plong District, Ngoc Linh Nature Reserve). – Laos. Known only from the southwest of the country: Xékong Province (Xe Sap National Biodiversity Conservation Area, Kaleum District; Phou Ajol, Dakchung District).
Biology. Stuart (2006) provided the first data on the ecology of this species. Parahelicops annamensis inhabits regions covered with tropical wet evergreen forest and subtropical montane evergreen forests between about 300 and 1,500 m a.s.l., although most specimens for which we have data were collected between 1,200 and 1,400 m a.s.l. All were collected in primary tropical or montane evergreen forests and in close association with fast-moving, rocky forest streams. This species seems to be mainly nocturnal and aquatic, although one specimen was found on the ground, under a log on the bank of a 4 m wide rocky stream. Specimen ZFMK 86457 was captured in the evening in the leaf litter of a forest stream at 510 m a.s.l., with only the snout tip protruding from the surface of the water. The diet and reproductive habits of Parahelicops annamensis are still unknown.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Parahelicops annamensis Bourret, 1934
| David, Patrick, Pauwels, Olivier S. G., Nguyen, Truong Quang & Vogel, Gernot 2015 |
Parahelicops annamensis
| Bourret 1934: 170 |