
Myrmecodesmus mundus ( Chamberlin, 1943 )
|
publication ID |
https://doi.org/ 10.5281/zenodo.155975 |
|
DOI |
https://doi.org/10.5281/zenodo.6279024 |
|
persistent identifier |
https://treatment.plazi.org/id/03E087E4-B11D-FF94-B36A-4C69FB8640EC |
|
treatment provided by |
Plazi |
|
scientific name |
Myrmecodesmus mundus ( Chamberlin, 1943 ) |
| status |
|
Myrmecodesmus mundus ( Chamberlin, 1943) View in CoL
Figs. 16
Eirenyma munda Chamberlin, 1943:67 View in CoL , pl. 16, figs. 162164. Loomis, 1968:49. Myrmecodesmus mundus: Shear, 1977:255 View in CoL . Hoffman, 1999:494.
Type specimens. Male holotype ( NMNH) collected by F. Bonet, 31 May 1941, at Veracruz, Veracruz state, Mexico. Two male and two female paratypes ( NMNH) collected by C. Bolivar and F. Bonet, 11 November 1941, at Atoyac, a town ca. 10 mi (16 km) E Cordoba or about 45 mi (72 km) W Veracruz City; coordinates for Atoyac are 96° 46' 10.8" W, 18°55' 13.5" N. Chamberlin (1943) stated that this sample contained six specimens, one male and five females, but today there are four, two males and two females.
Holotype. 20 segments including epiproct. Length ca. 3.4 mm, maximum width ca. 0.6 mm. Dorsum relatively light brown in color, paranota colorless; body heavily sclerotized. Collum with low, rounded, indistinct pustules, anterior margin relatively flat, not appreciably upturned, scallops shallowly separated, indistinct (Fig. 1). Tergite 2 with typical threelobed paranota (Fig. 2, top), paramedian tubercles coalesced into low, indistinct carinae, caudal segmental margin smooth and sublinear, slightly crenulated laterad. Remaining tergites more strongly crenulated laterad, carinae becoming progressively more distinct caudad, those on segment 19 moderately elongated into two broad lobes, overhanging and extending beyond epiproct, obscuring latter in dorsal view. Paranota with two indistinct, subequal lobes on segments 318, broadly rounded on 19 (Fig. 2); porosteles totally absent. Epiproct small and smoothly rounded, without lobes or crenulations. Legs and sterna without modifications.
FIGURES. 12. Myrmecodesmus mundus holotype. 1, profile of collum margin, dorsal view. 2, profiles of paranota of segments 3, 5, and 19.
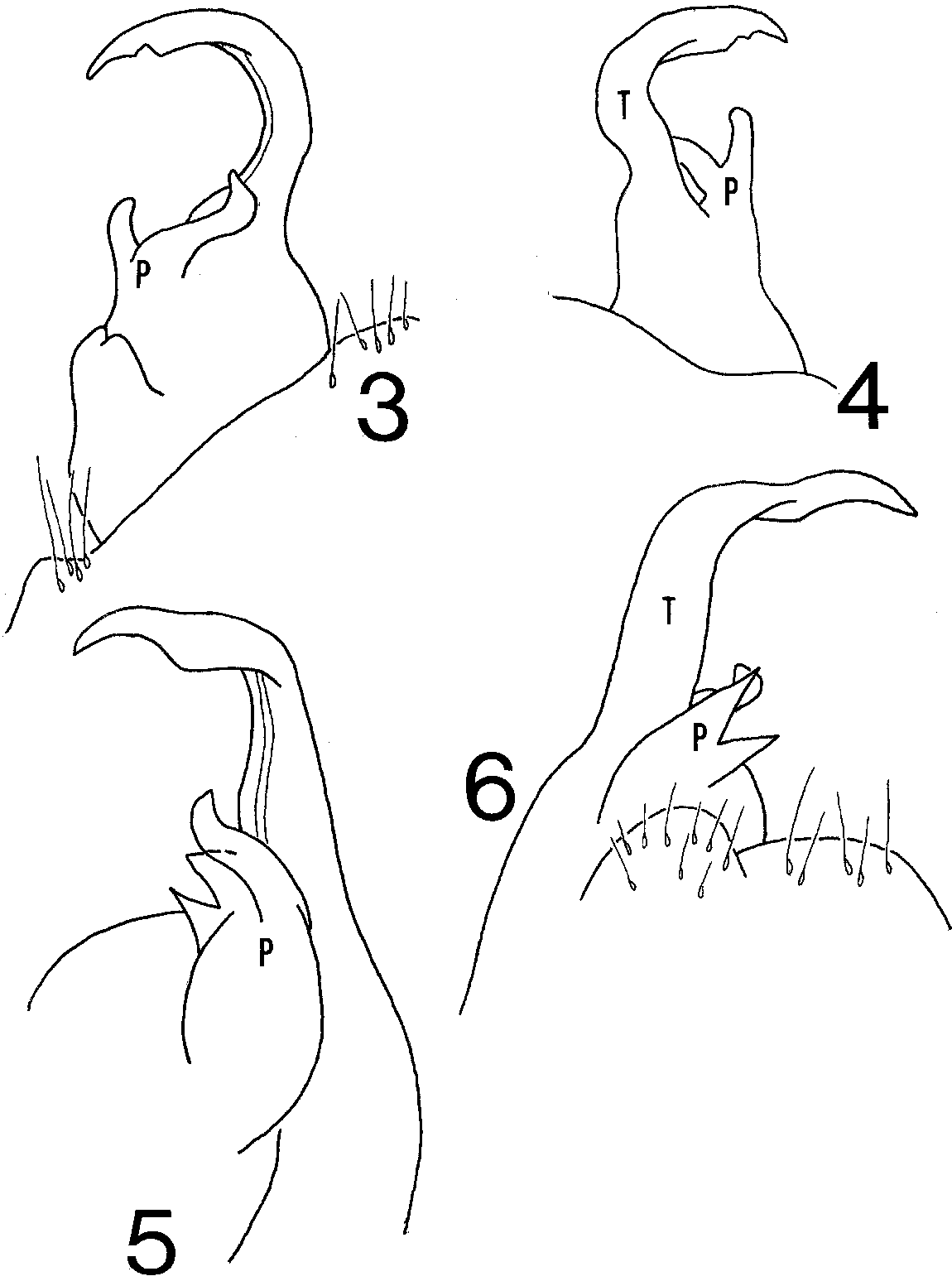
Gonopod ( Figs. 34 View FIGURES 3 6 ) minute, details visible only under high magnification; telopodite a simple, broadly curved blade, without branches, with slight flange distad on caudal side, tip curving slightly dorsad, acuminate. Prefemoral process short, divided into two branches, caudal one longer, closely appressed to base of telopodite, bending abruptly ventrad distally, narrowing apically; anterior branch short and broad, upright.
Male Paratype. Agreeing closely with holotype in somatic features but paranotal lobes slightly more distinct and caudally projecting crests on segment 19 shorter and narrower. Gonopod ( Figs. 56 View FIGURES 3 6 ) with telopodite upright basally, bending abruptly anteriad near midlength. Prefemoral process with caudal branch slightly sinuate but generally upright, anterior branch redivided into two sharply acuminate, subequal subbranches.
Ecology. The holotype was collected from under $leaves of Camarina#; the paratypes were encountered in $tropical woods under leaves.#
Distribution. Known only from the type and paratype localities.
Remarks. I cannot improve on Chamberlin s drawing (1943, fig. 163) of the caudal segments, but his illustration of the gonopodal telopodite of the holotype (1943, fig. 164), a simple, uncinate blade (a $hook#) without a prefemoral process, is wholly inadequate and misleading. While the telopodite lacks secondary projections, it possesses a distal flange on the caudal margin, and there is a minute prefemoral process consisting of two branches (figs. 34); the paratype prefemoral process consists of three branches, the anterior one redivided. Thus in this short distance of only 45 mi (72 km) there is seemingly significant variation in the structure of the prefemoral process, and the paratype s telopodite is more upright and bends abruptly, though still possessing a distocaudal flange. This difference may justify specific recognition of the paratypes, but more material and further study is needed to determine this definitely. However, the division of the prefemoral process becomes significant when it is noted that the structure in the Texas species, some 800 mi (1,280 km) to the north in Kendall County, is similarly divided into two ultra minute, acuminate banches ( Shelley 2003, figs. 1920), basically the configuration in the paratype of M. mundus minus the caudal branch. Likewise, the telopodite in the Texas species appears to be a simple, uncinate $hook# under low magnification, but high power reveals that it curves more strongly and is structurally complex, with a side branch, the presumed solenomere, and a divided and redivided tip. Consequently, the basic gonopodal pattern in both species is a simple, curvilinear, bladelike telopodite and divided prefemoral process. The affinities of the Texas species therefore appear to lie with M. mundus , and both are at the low end of the generic size spectrum, being about 3 mm in total length. Innumerable additional species of Myrmecodesmus probably await discovery in Texas and Mexico, so while we are likely only beginning to catalog the genus, we may be seeing the basis for delineating one $species group# that ranges at least from central Veracruz to the vicinity of San Antonio, Texas & minute forms with the aforementioned gonopodal pattern in which there may be telopodal elaborations and the caudal or possibly the anterior prefemoral branch may be lost. While the trend in diplopodology is not to recognize species groups taxonomically, it should be noted that the name, Eirenyma Chamberlin, 1943 , is available to denote a subgeneric entity if a future student decides such is warranted.
| NMNH |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Myrmecodesmus mundus ( Chamberlin, 1943 )
| Shelley, Rowland M. 2002 |
Eirenyma munda
| Hoffman 1999: 494 |
| Shear 1977: 255 |
| Loomis 1968: 49 |
| Chamberlin 1943: 67 |