
Sarcosuchus, SP. (DE BROIN & TAQUET, 1966)
|
publication ID |
https://doi.org/ 10.1093/zoolinnean/zlz057 |
|
DOI |
https://doi.org/10.5281/zenodo.5719172 |
|
persistent identifier |
https://treatment.plazi.org/id/03E11917-FFEE-FFE4-EDA4-FE7740A50267 |
|
treatment provided by |
Carolina |
|
scientific name |
Sarcosuchus |
| status |
|
SARCOSUCHUS SP. ( DE BROIN & TAQUET, 1966)
Referred material: A single broken osteoderm from the right side of the dorsal dermal shield ( BMNH R3224).
H o r i z o n a n d l o c a l i t y: S e t ú b a l l o c a l i t y, L o w e r Cretaceous Recôncavo Basin, Bahia Supergroup, Ilhas Group (Late Hauterivian–Early Barremian) with no more refinements.
TAXONOMIC DECISIONS
There is a common error in the literature about the type specimens of Sarcosuchus hartti . Some authors treat the specimen BMNH R3423 described by Mawson & Woodward (1907) as the holotype of this species (e.g. Andrade et al., 2011). Also, Norell & Storrs (1989) designated the specimen YPM 516 as its holotype, which is a single tooth said to be originally studied by Marsh (1869). However, those assignments are not in agreement with the rules of the ICZN (1999). In Marsh’s (1869) original description, he did not designate a holotype, nor did he provide the identity of the teeth he studied. He only mentioned the teeth illustrated by Allport (1860) and the teeth collected by Hartt. Therefore, the three teeth from Allport (1860, pl. XVI, figs 1–3, 5) plus the teeth collected by Hartt, must be treated as part of the type series of Sarcosuchus hartti . This is the case even with those teeth being lost at the present time, as recommended in articles 72.1.1 and 72.4 of the ICZN (1999). Based on the articles 72.2 and 74 of the ICZN (1999), we propose that the specimen YPM 516 should not be treated as the holotype, but as a lectotype of Sarcosuchus hartti . This is justified, because the specimen belongs to the type series of Marsh (1869) and was designated as type material by Norell & Storrs (1989). Also, the remaining specimens of the type series (e.g. Allport’s specimens) must be treated as paralectotypes based on articles 72.1.3 and 74.1.3 of the ICZN (1999). The referral of the specimens described by Mawson & Woodward (1907) to Sarcosuchus hartti , which were latter discussed by Buffetaut & Taquet (1977), should be regarded with caution, being recognized as referred materials only.
The large tooth BMNH R2983 ( Fig. 3B, C View Figure 3 ), described by Mawson & Woodward (1907), was also referred to Sarcosuchus hartti . However, this material has lost all the enamel and, therefore, cannot be properly assignated to this species due to the absence of autapomorphic features; but, based on the overall morphology, this specimen can be considered a crocodyliform.
The discovery of the mandible (BMNH R3423) with in situ teeth presenting ‘delicate wrinkled surface’ suggests that the teeth from Bahia State (Recôncavo Basin) belong to Sarcosuchus hartti . Even though the specimen BMNH R3224 (an osteoderm) came from the same locality as the mandible (BMNH R3423), Mawson & Woodward (1907) gave no detailed taphonomic information about this specimen, thus the assignment of the osteoderm to the same species of the other material from Bahia (both the mandible and teeth) is hampered. Therefore, based on the morphological features properly described below, the specimen BMNH R3224 can only be assigned to Sarcosuchus sp.
The final Brazilian material referred to Sarcosuchus is a procoelous centrum of a dorsal vertebra collected and classified by Owen, which described the reptilian material in Allport (1860), as a ‘dinosaurian reptile’ with affinities to Megalosaurus . Mawson & Woodward (1907) described this material as Megalosauridae . Later it was referred to a ‘carnosaur’ dinosaur by Buffetaut & Taquet (1977). More recent works suggested, first, that this vertebra could belong to Sarcosuchus hartti ( Campos & Kellner, 1991) , and was further assigned as an indeterminate crocodyliform ( Kellner & Campos, 2000). Unfortunately, this specimen is lost and it is, therefore, not certain which crocodyliform species it represents.
FULL DESCRIPTION
Dentition
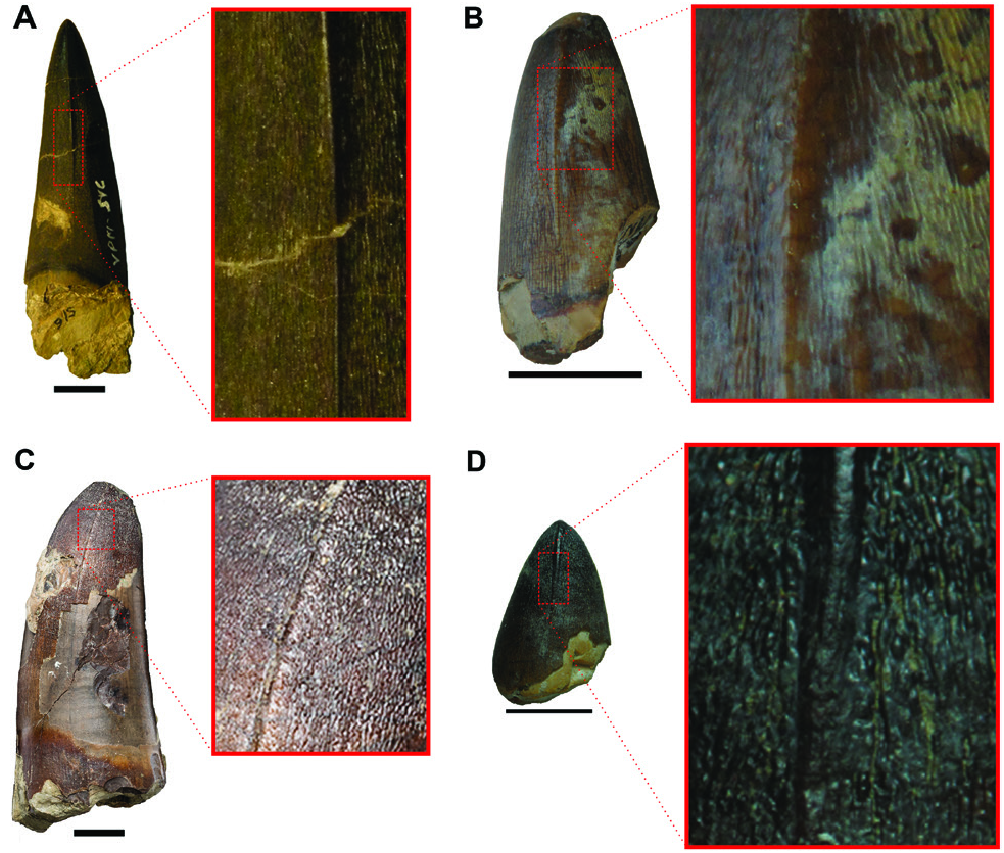
The teeth illustrated by Allport (1860: pl. XVI, figs 1–3, 5) are described as having a ‘delicate wrinkled surface’ (not the same used for Theropoda teeth, see: Brusatte et al., 2007). Marsh (1869) complements that description, stating that they show cutting edges and refines Allport’s (1860) definition of wrinkles as a peculiar striate pattern that was not seen in other specimens before.
YPM 516 is 70 mm in height with the crown alone comprising 60 mm ( Fig. 3A View Figure 3 ). Only a small portion of the root is preserved. The tooth is almost straight with a very gentle curvature to the lingual face. The enamel presents many delicate longitudinal and oblique lines forming an anastomosing surface along the entire crown ( Fig. 4A View Figure 4 ). The anterior and posterior carinae are composed of well-marked longitudinal ridges with smooth crenulations on them ( Figs 3A View Figure 3 , 4A View Figure 4 ). Therefore, this tooth can be described as false-ziphodont in morphology (sensu Prasad & de Broin, 2002). The carinae do not touch the basal margin of the tooth, presenting a progressive growing until the apex ( Fig. 3A View Figure 3 ). The tooth is subcircular in cross-section. Based on the general morphology of Sarcosuchus imperator, YPM 516 is probably a ‘caniniform’ anterior tooth (sensu D’ Amore et al., 2019) in having a straight, tall and sharp crown.
Here we present a more refined description of the paralectotypes based on Allport’s (1860: pl. XVI, figs 1–3, 5) illustrations. The teeth illustrated in Allport’s figures 1, 2, 3 and 5, are referred to here as teeth one, two, three and four, respectively. Tooth one presents most of the characteristics described for YPM 516. It differs only in the longitudinal ridge on the carinae, which goes all the way long from the base to the apex of tooth one. Tooth two is apicobasally short, being smaller and stouter in comparison with YPM 516 and tooth one. The enamel is badly preserved, being restricted to the extreme convex apex. The labial face is slightly curved lingually, while the lingual face is straight. The enamel anastomosing surface is inferred based on Allport’s (1860) classification of teeth morphotypes. Tooth three has a well-preserved anastomosing enamel surface. Like tooth two, it is also apicobasally short, but tooth three differs from tooth four in having a more acute apex, the result of the stronger curvature on the lingual surface. Tooth four slightly resembles YPM 516 and tooth one, but it differs in being less robust and presenting a sigmoid curvature in the labial face. The illustration just enables us to infer a subcircular cross-section for teeth two, three and four. Also, the longitudinal ridges on the carinae can only be observed in tooth one.
The two teeth studied by Mawson & Woodward (1907) and Buffetaut & Taquet (1977) are BMNH R2983 ( Fig. 3B, C View Figure 3 ) and BMNH R3079 ( Fig. 3D, E View Figure 3 ). The specimen BMNH R 2983 is 54 mm in height, with a straight lingual margin and a slightly curved labial margin ( Fig. 3B, C View Figure 3 ). It is subcircular in cross-section with a major axis of 33 mm. The enamel is badly preserved and does not show the longitudinal and oblique lines, but some deep, parallel scratches are present on the surface of the tooth ( Fig. 3B, C View Figure 3 ). The carinae are also not preserved and the apex presents a well-marked wear facet ( Fig. 3B, C View Figure 3 ). Due to the absence of the diagnostic features, this tooth cannot be assigned to Sarcosuchus hartti . BMNH R3079 is 88 mm in height, with both lingual and labial margins slightly curved lingually ( Fig. 3D, E View Figure 3 ). This tooth shows a subcircular cross-section with a major length of 40 mm. The enamel is partially preserved, mainly at the apex and on the lingual margin. It presents the characteristic delicate longitudinal striation that forms the anastomosing surface of the tooth ( Fig. 4C View Figure 4 ). This pattern is particularly well-preserved at the tooth apex. The longitudinal ridges on the carinae are preserved on both sides of the apex, being slightly developed. The apex is concave without any wear facets.
The specimen MN 7460-V is 25 mm in height ( Fig. 3F, G View Figure 3 ). The labial margin is slightly convex and the lingual margin is gently concave, thus the apex assumes a more lingual-oriented position ( Fig. 3F, G View Figure 3 ). The enamel shows the characteristic delicate longitudinal and oblique lines that form the anastomosed surface ( Fig. 4B View Figure 4 ). The tooth apex is concave and the enamel was lost, what could be a wear facet ( Fig. 3F, G View Figure 3 ). The crosssection is circular. Both carinae have well-marked longitudinal ridges, being completely absent next to the base ( Fig. 3F, G View Figure 3 ). The specimen MN 7461-V ( Fig. 3H, I View Figure 3 ) is shorter in height than the first one, with a total length of 22 mm. Both lingual and labial facets are convex with a more rounded than pointed apex (‘molariform’ sensu D’ Amore et al., 2019; Fig. 3H, I View Figure 3 ). The cross-section is circular. The enamel and the carinae have the same characteristics observed in MN 7460-V ( Fig. 4D View Figure 4 ).
Mandible
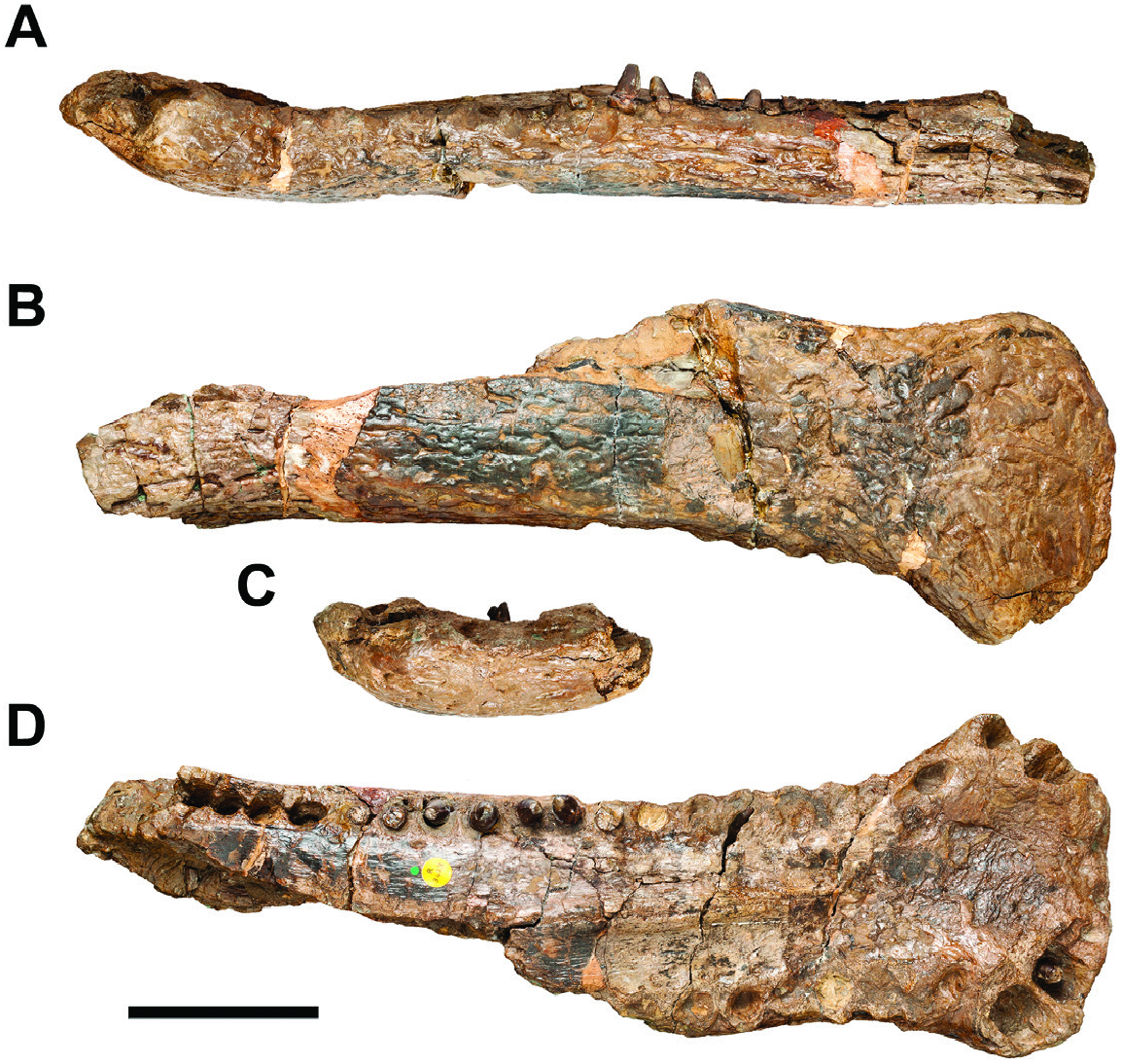
In the anterior portion, specimen BMNH R3423 consists of an incomplete mandible ( Figs 5 View Figure 5 , 6 View Figure 6 ). The dentary is only preserved bone, but it is possible to observe the region where the splenial would contact it. The maximum preserved length of the mandible is 430 mm, but the total length of the lower jaw could reach at least twice this length. The mandibular symphysis is elongated ( Figs 5B, D View Figure 5 , 6B, D View Figure 6 ) and dorsoventrally flattened ( Figs 5A, C View Figure 5 , 6A, C View Figure 6 ). The anteriormost region is spatulated in shape due to the lateral expansion of the dentaries ( Figs 5D View Figure 5 , 6D View Figure 6 ). It flares up at the level of the fifth alveolus and reaches its maximum width at the level of the fourth alveolus ( Figs 5D View Figure 5 , 6D View Figure 6 ). In this region, the dorsal surface has a shallow concavity, which is more rugose than the remaining surface of the mandible ( Figs 5D View Figure 5 , 6D View Figure 6 ). The dorsal surface going from the region next to the final portion of the anterior splenial process to the eighth alveolus is rugose ( Figs 5D View Figure 5 , 6D View Figure 6 ). The dorsal surface posterior to the anterior flared region shows a low median sagittal keel, with the syphysis between the dentaries comprising its apex ( Figs 5D View Figure 5 , 6D View Figure 6 ). There are depressions on each side of the sagittal keel, between the midline and the medial alveolar margins ( Fig. 5D View Figure 5 ). There is no participation of the splenials in the medial margins of the posterior alveoli that are preserved ( Fig. 5D View Figure 5 ). The ventral surface of the dentary has a coarse ornamentation with shallow interconnected grooves ( Fig. 5B View Figure 5 ). In lateral view, the mandible shows a linear ventral margin up to the posterior border of the fourth alveolus and from this point beyond it slopes anterodorsally ( Figs 5A View Figure 5 , 6A View Figure 6 ). The laterodorsal profile of the mandible is double festooned (i.e. two distinct concave regions), the first and deeper concavity is located between the fourth and tenth alveoli, while the second one is shallow and is placed between the 13 th and the 21 st alveoli ( Figs 5A View Figure 5 , 6A View Figure 6 ). There are 22 dentary alveoli on the left side of the mandible and 11 on the right side. The first two pairs of teeth are transversally aligned in the anterior margin of the dentary ( Figs 5D View Figure 5 , 6D View Figure 6 ). They are procumbent and smaller in comparison with all other teeth ( Figs 5D View Figure 5 , 6D View Figure 6 ). The third and fourth pair of alveoli are similar in size, but the fourth ones are the largest teeth in the dentary ( Figs 5D View Figure 5 , 6D View Figure 6 ). These teeth are lateroposteriorly tilted relative to the second pair of alveoli, being the third pair more medially than the fourth ( Figs 5D View Figure 5 , 6D View Figure 6 ). There is a diastema between the fourth and fifth alveoli, being the diastema similar in width to the size of the fourth alveoli ( Figs 5D View Figure 5 , 6D View Figure 6 ). A depressed region is present close to the diastema and probably represents a notch for an enlarged tooth at the premaxilla–maxilla ( Figs 5D View Figure 5 , 6D View Figure 6 ). Starting from the fifth pair of alveoli, all others are anteroposteriorly aligned ( Figs 5D View Figure 5 , 6D View Figure 6 ). They are similar in size, but show different shapes from subcircular to labiolingually elliptic outlines ( Figs 5D View Figure 5 , 6D View Figure 6 ). The posterior series of teeth are packed together, separated by thin bone bars between each alveolus ( Figs 5D View Figure 5 , 6D View Figure 6 ). The only exceptions are the last four sockets, which are confluent and form a dental groove ( Figs 5D View Figure 5 , 6D View Figure 6 ).
Specimen BMNH R3423 shows seven complete teeth preserved in situ ( Fig. 5A, D View Figure 5 ), all of them present the autapomorphic ornamentation of the enamel, allowing us to assign this material to Sarcosuchus hartti . There is one ‘caniniform’ tooth partially erupted on the third alveolus of the right dentary, which is similar in morphology to the lectotype specimen YPM 516. The remaining teeth are on the left side of the dentary from the 12 th to the 17 th, and the morphology exhibited by them is like those observed in MN 7460-V and MN 7461-V. Therefore, the dentition has a heterodont pattern, with at least the third teeth being a tall caniniform, while the posterior ones are blunter and shorter. The mandible shows occlusion marks ( Fig. 5D View Figure 5 ), with the anterior ones visible in lateral view and the posterior ones in dorsal view, near the lateral margins of the alveoli. The distribution pattern of those toothmarks indicates overbite occlusion.
The new specimen MN 7459-V consists of a fragment of the right ramus of the mandibular symphysis ( Fig. 7 View Figure 7 ). There are 14 preserved alveoli and two associated teeth (specimens MN 7460-V and MN 7461- V). The alveoli are similar in size with two different outlines: subcircular and labiolingually elliptical ( Fig. 7A View Figure 7 ). The interalveolar space is smaller than the anteroposterior length of the alveoli ( Fig. 7A View Figure 7 ). There are several tooth marks in the interalveolar spaces posterior to the seventh alveolus ( Fig. 7A View Figure 7 ). The first two marks are placed near the lateral margin of the alveoli and the posterior ones are displaced medially in relation with each other. The last toothmark is preserved near the medial margin of the bone, immediately posterior to the last preserved alveolus. There are also some small aligned foramina close to the medial margin of the alveoli ( Fig. 7A View Figure 7 ). The ornamentation in the ventral surface is more tenuous and with more sparse pits than that observed in BMNH R3423 ( Fig. 7B, D View Figure 7 ). In dorsal view, it is possible to see the sutural region between the splenial and the dentary ( Fig. 7A View Figure 7 ). Also, it is possible to observe the dentary–dentary symphysis in medial view ( Fig. 7C View Figure 7 ) and the dentary–splenial contact posteriorly, with a concavity between the dorsal and ventral articular regions ( Fig. 7C View Figure 7 ). The posterior portion of the dentary flares laterally to receive the anterior process of the splenial that wedges anteriorly to the level of the sixth alveolus ( Fig. 7A View Figure 7 ). A low median sagittal keel is observed in the mid-palatal region of the mandible, where the dentaries contact each other ( Fig. 7A View Figure 7 ). Located between this keel and the medial alveolar margin there is a rugs area that ends near the anterior process of the splenial ( Fig. 7A View Figure 7 ).
Osteoderm
The specimen BMNH R3224 is a partially preserved osteoderm ( Fig. 8 View Figure 8 ). It is subrectangular in shape and has a straight, smooth and vertical anterior margin for articulation with the anterior osteoderm ( Fig. 8A View Figure 8 ). There is a short and robust anterior process, which is placed at the lateral margin of the osteoderm ( Fig. 8A View Figure 8 ). The bony process is smooth on its surface like the anterior and lateral margins and is slightly directed medially ( Fig. 8A View Figure 8 ). There is a low keel separating the large dorsomedial surface of the osteoderm from the smaller lateral region ( Fig. 8A View Figure 8 ). The keel runs obliquely following the orientation of the anterior bony process. The lateral area is convex and follows the same orientation of the anterior process ( Fig. 8A View Figure 8 ). Because of such orientation, it is possible to infer that the posterior margin was wider than the anterior one, giving a trapezoidal shape for the complete osteoderm. The dorsal surface of the osteoderm is ornamented with a great number of large, deep and isolated pits that varies in shape ( Fig. 8A View Figure 8 ). The ventral facet is smooth with no muscular scars ( Fig. 8B View Figure 8 ). The osteoderm is thick in lateral view proportionally to the other surfaces ( Fig. 8B View Figure 8 ). The broken region allows the observation of two different kinds of bone tissue; a more compact bone that composes the dorsal, ventral and lateral regions of the osteoderm, and a spongy tissue located in the inner region ( Fig. 8B View Figure 8 ). Based on comparisons with the specimen MNN 607 of Sarcosuchus imperator , it is possible to suggest that this osteoderm would be a more anterior element of the dermal shield.
PHYLOGENY
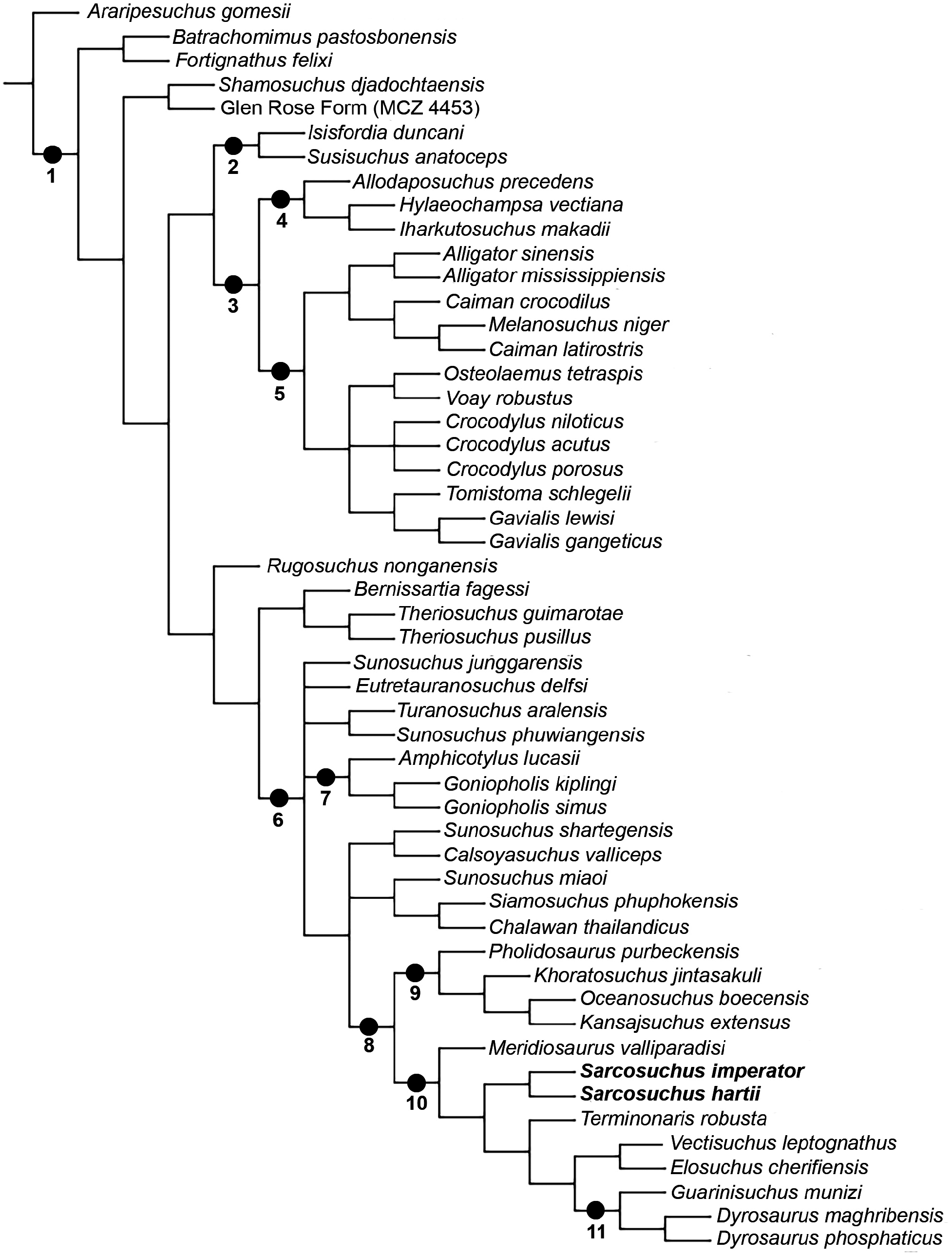
The phylogenetic analysis resulted in 12 minimumlength trees (see Supporting Information, Supplementary Information X for the graphical representation of those trees), with 1287 steps (CI: 0.327 and RI: 0.610). The topological hypotheses are summarized and presented on the resulting strict consensus tree ( Fig. 9 View Figure 9 ).
The first lineage of Neosuchia , sister to all other species, is the clade uniting Batrachomimus and Fortignathus . This relationship is supported by the share of an elongated symphyseal region, which is based on the proportion between length and width (Char. 222: 1 -> 2; i.e. the ancestor species has the condition 1, while the descendent species hereditary acquired the modified condition 2). Its sister-clade is formed by the remaining neosuchian species, and is supported by the following synapomorphies: ornamented region below the dental groove of the dentary (Char. 18: 1 -> 0); anteriormost end of the frontal acute (Char. 98: 0 -> 1); presence of the spina quadratojugalis at the posterior margin of the lower temporal fenestra (Char. 145: 0 -> 1); flat symphyseal region at the dorsal surface of mandible (Char. 221: 1 -> 0); and an aligned occlusion pattern of the premaxilla–dentary (Char. 246: 1 -> 0). Also, in some of the minimum length trees, the lateral margin of the upper temporal bar, postorbital–squamosal, being convex (Char. 128: 1 -> 0) is recovered as a synapomorphy.
The clade (( Susisuchidae + Eusuchia) ( Rugosuchus (( Bernissartia (Theriosuchus )) (Coelognathosuchia))) is supported by the following synapomorphies: external nares opens dorsally (Char. 40: 0 -> 3); double festooning at the lateroventral margin of the maxilla (Char. 64: 2 -> 3); the quadrate condyles are at the same level as the occipital condyle (Char. 155: 0 -> 1); rounded choana (Char. 187: 3 -> 1); laminar neural spines at the posterior series of cervical vertebrae (Char. 318: 1 -> 0); and absence of a rounded fossa between the base of the neural spine and the postzygapophyses in dorsal vertebrae (Char. 325: 1 -> 0).
The clade composed by ( Susisuchidae + Eusuchia) is supported by the following synapomorphies: absence of a perinarial fossa in the premaxilla (Char. 49: 1-> 0); absence of a notch in the lateral surface of premaxilla– maxilla contact (Char. 59: 1 -> 0); premaxilla–maxilla contact with sinusoid orientation at the palatal surface (Char. 61: 0 -> 1); a narrow rectangular frontal (Char. 97: 01 -> 2); a subequal proportion between anteroposterior and lateromedial axes of choana (Char. 188: 0 -> 1); anterior margin of choana posterior to the suborbital fenestrae (Char. 192: 1 -> 2); choana completely enclosed within the pterygoid (Char. 208: 12 -> 3); the proximal end of the scapular blade is wider than the distal end (Char. 291: 1 -> 0); presence of a notch at the dorsal margin of the iliac blade (Char. 307: 0 -> 1); procoelous centra in cervical vertebrae (Char. 320: 0 -> 1); biconvex articulation in the first caudal vertebrae (Char. 333: 0 -> 1); procoelous centra in caudal vertebrae (Char. 334: 0 -> 1); and the cervical osteoderms differs morphologically from the dorsal ones (Char. 346: 0 -> 1).
The sister-group of ( Susisuchidae + Eusuchia) is the clade ( Rugosuchus (( Bernissartia (Theriosuchus )) (Coelognathosuchia)) , which is supported by three synapomorphies: absence of occipital exposition of the cranioquadrate passage (Char. 122: 0 -> 1); absence of supraoccipital exposure at the skull roof (Char. 162: 1 -> 0); and choana anteriorly positioned in relation to the anterior margin of the pterygoid (Char. 193: 1 -> 0).
A shared recent ancestrality between Tomistoma and Gavialis is not supported here by any unambiguous character transformation. The phylogenetic position of true-gharials ( Gavialis ) and false-gharial ( Tomistoma ) have been a subject of continuous scientific debate for almost forty years, and it is now a classic case of phylogenetic conflict in the literature (Buffetaut, 1982, 1985; Norell 1989; Brochu 1997; Gatesy et al. 2003; Piras et al. 2010; Iijima &, Kobayashi, 2019). Usually molecular analyses consider both lineages as sister taxa diverging each other during Eocene or mid-Miocene, whereas morphology-based analyses set Gavialis apart from Tomistoma and other crocodylian species, and the stratigraphic data imply that the split of the Gavialoidea lineage occurred by the Late Cretaceous. The oldest gavialoid remains are from Upper Cretaceous and Paleocene deposits of North America, Europe and Africa ( Hua & Jouve, 2004). Today, the critically endangered Gavialis is limited to the northern part of the Indian subcontinent. However, the zenith of gavialoid diversity is in the South American Miocene (Riff et al. 2010; Scheyer et al., 2013; Cidade et al. 2019). Contradicting the results presented here in this respect, previous studies including the morphologically varied South American Neogene gavialoids strongly supported the monophyly of Gavialoidea and its basal-most position within Crocodylia (e.g. Brochu & Rincón 2004; Vélez-Juarbe et al. 2007; Riff & Aguilera, 2008; Bona et al., 2017; Salas-Gismondi et al. 2017).
Clade (( Bernissartia (Therioschus )) (Coelognathosuchia)) is supported by: the external nares are anterodorsal oriented (Char. 40: 3 -> 2); the anterior and posterior processes of jugal are subequal (Char. 133: 2 -> 1); triangular choana (Char. 187: 1 -> 2); dentary tooth medially located in relation to the premaxilla–maxilla contact (Char. 282: 0 -> 1); and dorsal osteoderms wider than long (Char. 337: 1 -> 2). In some minimum-length trees, the squamosal posterior processes are parallel (Char. 113: 1 -> 0).
The clade Coelognathosuchia shows a polytomy of several smaller clades (( Sunosuchus junggarensis ) ( Eutretauranosuchus ) ( Turanosuchus + Sunosuchus phuwiangensis ) ( Amphicotylus + Goniopholis ) (node 64)). Coelognathosuchia itself is well supported by the following synapomorphies: nasal does not participate in the borders of the external nares (Char. 76: 0 -> 1); the frontal is wide in the region between the orbits (Char. 95: 0 -> 1); the frontal posterior margin is positioned at the postorbital bar in the skull roof (Char. 103: 1 -> 0); the quadrate posterior ramus are at the same level of the maxillary teeth row (Char. 154: 2 -> 1); elliptic mandibular fenestra (Char. 214: 1 -> 0); the posterior teeth of maxilla–dentary are circular in cross-section (Char. 265: 0 -> 1); dentary alveoli 3 and 4 are close to each other (Char. 277: 0 -> 1); the third dentary alveoli are anterior to the fourth ones (Char. 279: 1 -> 2); and absence of a longitudinal keel at dorsal osteoderms (Char. 340: 0 -> 1). Some of the minimum-length trees present the following features as synapomorphies: presence of a non-ornamented region below the dental groove of dentary (Char. 18: 0 -> 1); lacrimal present a long length in relation to the width (Char. 84: 0 -> 1); robust postorbital bar (Char. 131: 0 -> 1); and robust splenial posterior to the mandibular symphysis (Char. 238: 0 -> 1).
The polytomy results from two diverging hypotheses found in the minimum-length trees.The first hypothesis is observed on minimum-length trees 1, 9, 10 and 11 and consists of (( Eutretauranosuchus ( Amphicotylus + Goniopholis )) ( Sunosuchus junggarensis (( Turanosuchus + Sunosuchus phuwiangensis ) (node 64)))).
Clade (( Eutretauranosuchus ( Amphicotylus + Goniopholis )) is supported by all minimum lengthtrees by follow features: the anteromedial region of the orbits present a transversal crest (Char. 8: 0 -> 1); there is no ornamentation in the surface below the dental groove (Char. 18: 0 -> 1); the posterior margin of the nasals are between the anterior margins of the orbits (Char. 80: 0 -> 1); the squamosal present an elongated posterior process (Char. 112: 0 -> 1); the temporo-orbital opening does not expose dorsally (Char. 115: 0 -> 1); the mandibular fenestra is horizontal at its anteroposterior axis (Char. 215: 1 -> 0); and the last premaxillary tooth is anteromedial in relation with the first maxillary tooth (Char. 256: 1 -> 2).
Clade ( Amphicotylus + Goniopholis ) accounts for the following characters: perinarinal crest present in the premaxilla (Char. 1: 0 -> 1); preorbital crest present in the anterior region of the orbits (Char. 5: 0 -> 1); the postnarial fossa on premaxilla is present (Char. 51: 0 -> 1); the premaxilla has a hatchet-like shape in dorsal view (Char. 55: 0 -> 4); there are paired maxillary depressions at the laterodorsal surface of maxilla (Char. 71: 0 -> 1); the posterior ramus of prefrontal is long reaching the median region of the orbits (Char. 90: 0 -> 1); the frontal is wide (Char. 94: 0 -> 1); the frontal has two unlevelled surfaces (Char. 99: 0 -> 1); and, at the skull table the parietal-squamosal suture is grooved (Char. 108: 0 -> 1).
The ( Sunosuchus junggarensis (( Turanosuchus + Sunosuchus phuwiangensis ) (node 64))) lineage has some particular features recovered in all minimum length-trees, which are: small anterior palpebrals (Char. 35: 1 -> 0); the incisive foramen is small (Char. 45: 0 -> 1); premaxilla has a paddle-like shape in dorsal view (Char. 55: 0 -> 3); and there are vascular openings at the dorsolateral surface of posterorbital bar (Char. 126: 0 -> 1).
The node (( Turanosuchus + Sunosuchus phuwiangensis ) (node 64))) is supported by only one character that is the premaxilla overbiting the mandible in occlusion (Char. 246: 0 -> 1). While ( Turanosuchus + Sunosuchus phuwiangensis ) has the following synapomorphies: wedge-shaped mandible (Char. 211: 1 -> 0); the anterior region of the symphyseal portion, in lateral view, is acute (Char. 216: 1 -> 0); and the symphyseal region has the anterior width smaller than the posterior (Char. 218: 2 -> 0).
The second hypothesis is observed in minimumlength trees 2, 3, 4, 5, 6, 7, 8 and 12, and consists of (( Amphicotylus + Goniopholis ) (( Eutretauranosuchus ( Sunosuchus junggarensis ( Turanosuchus + Sunosuchus phuwiangensis ))) (node 64))). The ( Amphicotylus + Goniopholis ) is well supported by several features in all minimum length-trees, which are: perinarinal crests present in external nares (Char. 1: 0 -> 1); preorbital crest present (Char. 5: 0 -> 1); there is a transversal crest in the anteromedial region to the orbits (Char. 8: 0 -> 1); there is a postnarial fossa (Char. 51: 0 -> 1); there are paired maxillary depressions in the laterodorsal surface of maxilla (Char. 71: 0 -> 1); the prefrontal posterior ramus is long (Char. 90: 0 -> 1); the region between orbits is wide (Char. 94: 0 -> 1); the frontal has two unlevelled surfaces (Char. 99: 0 -> 1); the parietal–squamosal extends over the occipital surface (Char. 108: 0 -> 1); the temporoorbital opening is not exposed dorsally (Char. 115: 0 -> 1); the maxillary teeth row is in the same level of the occipital condyle (Char. 178: 0 -> 1); the last premaxillary tooth is anteromedial located in relation to the first maxillary tooth (Char. 256: 1 -> 2); there is a paradistal rotation of the dentition in relation to the medial sagittal axis (Char. 262: 0 -> 1); and the crown of anterior dentition is bulbous (Char. 270: 0 -> 1).
Clade(( Eutretauranosuchus (Sunosuchusjunggarensis ( Turanosuchus + Sunosuchus phuwiangensis ))) (node 64))) is supported by: the anterior palpebrals are small in size (Char. 35: 1 -> 0); the incisive foramen is small (Char. 45: 0 -> 1); in the lateral surface of the premaxilla– maxilla there is an indentation (Char. 60: 1 -> 0); there is a fossa close to the alveolar margin in the maxilla (Char. 70: 0 -> 1); the premaxilla overbites the dentary (Char. 246: 0 -> 1); and the first two alveoli pairs from mandible are smaller than the third and fourth ones (Char. 278: 2 -> 0).
The clade ( Eutretauranosuchus ( Sunosuchus junggarensis ( Turanosuchus + Sunosuchus phuwiangensis ))) has the following characters supporting it: there is a single festooning at the lateroventral margin of maxilla (Char. 64: 3 -> 2); the maxilla–prefrontal suture is present (Char. 73: 0 -> 1); and the first pair of dentary alveoli are anterior in relation to alveoli three and four (Char. 275: 0 -> 1). The inner clade ( Sunosuchus junggarensis ( Turanosuchus + Sunosuchus phuwiangensis )) accounts for only one character: that is the surface below the dental groove is ornamented (Char. 18: 1 -> 0). The ( Turanosuchus + Sunosuchus phuwiangensis ) clade is supported by: wedge-shaped mandible (Char. 211: 1 -> 0); the anterior width of the symphyseal region is smaller in relation to the posterior one (Char. 218: 2 -> 0); and there is a hypertrophied tooth adjacent to premaxilla–maxilla suture (Char. 281: 0 -> 1).
The polytomy (( Sunosuchus shartegensis + Calsoyasuchus ) ( Sunosuchus miaoi ( Siamosuchus + Chalawan ))) ( Tethysuchia )), node 64 in the consensus tree (see Supporting Information, Supplementary Information X), is the result of two competing topological hypotheses present in the minimum-length trees. This polytomy is supported by the following features as synapomorphies: the surface below the dental groove is non-ornamented (Char. 18: 0 -> 1); the posterodorsal process of the premaxilla extends posterior to the third maxillary alveolus (Char. 53: 0 -> 1); there is no spina quadratojugalis in the lower temporal fenestra (Char. 145: 1 -> 0); there are lateral openings to the main Eustachian tube (Char. 159: 0 -> 1); the choana is rectangular (Char. 187: 2 -> 3); the symphyseal region is extremely long (Char. 222: 2 -> 3); the splenial has a long participation in the mandibular symphysis (Char. 237: 1 -> 2); and there are no hypertrophied teeth in the anterior dentition of the maxilla (Char. 257: 0 -> 1).
The first competing hypothesis of the aforementioned polytomy is observed in minimum-length trees 1, 3, 5, 6, 7, 9 and 11 and consists of (( Sunosuchus miaoi ( Siamosuchus + Chalawan )) (( Sunosuchus shartegensis + Calsoyasuchus ) ( Tethysuchia ))). The clade ( Sunosuchus miaoi ( Siamosuchus + Chalawan )) has the following features to support it: the preorbital bones are elevated laterodorsally (Char. 26: 0 -> 1); the surangular participates in the glenoid fossa (Char. 228: 0 -> 1); and the surangular extends to the posterior region of the retroarticular process (Char. 232: 0 -> 1). The ( Siamosuchus + Chalawan ) clade is supported by two features: the quadrate medial condyle is hypertrophied (Char. 156: 0 -> 1); and supraoccipital exposes on skull roof (Char. 162: 0 -> 1). The clade ( Sunosuchus shartegensis + Calsoyasuchus ) ( Tethysuchia )) accounts for: the premaxilla does not taper between maxilla and nasal (Char. 52: 1 -> 0); the maximum width of the premaxilla is wider than the maxilla (Char. 54: 0 -> 1); the external auditory meatus is large (Char. 117: 1 -> 0); and the mandible symphyseal region is parallel to the horizontal plane (Char. 217: 0 -> 1). Finally, the clade ( Sunosuchus shartegensis + Calsoyasuchus ) is supported by: the narrow platyrostral rostrum (Char. 30: 4 -> 3); the snout has an external antorbital fenestra (Char. 43: 0 -> 1); and, the premaxilla–maxilla suture is sinusoid in palatal surface (Char. 61: 0 -> 1).
The second hypothesis is based on the topological relationship observed in the minimum-length trees 2, 4, 8, 10 and 12, which is ((( Sunosuchus shartegensis + Calsoyasuchus ) ( Sunosuchus miaoi ( Siamosuchus + Chalawan ))) ( Tethysuchia )). The (( Sunosuchus shartegensis + Calsoyasuchus ) ( Sunosuchus miaoi ( Siamosuchus + Chalawan ))) lineage accounts for: the absence of incisive foramen (Char. 44: 0 -> 1); the frontal–parietal suture rests on skull roof (Char. 104: 2 -> 0); the mandibular fenestra is large in relation to the orbit (Char. 213: 0 -> 1); and the mandibular fenestra is oblique at its anteroposterior axis (Char. 215: 0 -> 1). The ( Sunosuchus shartegensis + Calsoyasuchus ) clade is supported by two characters: a narrow platyrostral rostrum (Char. 30: 4 -> 3); and the snout present an external antorbital fenestra (Char. 43: 0 -> 1). The clade ( Sunosuchus miaoi ( Siamosuchus + Chalawan )) is supported by: the periorbital bones are laterodorsally elevated (Char. 26: 0 -> 1); the maxillopalatal fenestra is present (Char. 74: 0 -> 1); the postorbital bar is thin (Char. 131: 1 -> 0); the surangular participates on the glenoid fossa (Char. 228: 0 -> 1); and the surangular extend to the posterior region of the retroarticular process (Char. 232: 0 -> 1). Finally, the ( Siamosuchus + Chalawan ) clade is united by two characters: the quadrate medial condyle is hypertrophied (Char. 156: 0 -> 1) and the supraoccipital is exposed on the skull roof (Char. 162: 0 -> 1).
The Tethysuchia is supported in all minimum length-trees by the following characters: the premaxilla ventral margin is ventral in relation to the maxillary ventral margin (Char. 56: 1 -> 2); the maxilla lateroventral margin is straight (Char. 64: 3 -> 0); the lateral border posterior to the external narial opening of nasal is oblique to the sagittal axis (Char. 78: 1 -> 0); the postorbital bar is elliptic in cross-section (Char. 130: 1 -> 0); and the dentary does not extend below the mandibular fenestra (Char. 223: 1 -> 0). In some trees it is also united by: there being a posteroventral process of lacrimal beneath the orbits (Char. 85: 0 -> 1).
Within Tethysuchia there is a new clade Tethysuchoidea, which consists of ( Meridiosaurus (( Sarcosuchus ) ( Terminonaris (( Elosuchus + Vectisuchus ) ( Dyrosauridae )))), being supported by: posterodorsal process of premaxilla extends anterior to the third maxillary alveolous (Char. 53: 1 -> 0); the ventral margin of premaxilla and maxilla are at the same level (Char. 56: 2 -> 1); occlusion pits for the first dentary teeth at the palatal surface of premaxilla (Char. 58: 0 -> 1); no lacrimal process beneath the orbits (Char. 85: 1 -> 0); upper temporal bar is oblique (Char. 127: 0 -> 1); pterygoid participates at the posterior, lateral and parts of the anterior margin of the choana (Char. 208: 1 -> 2); and posterodorsaldirected retroarticular process in lateral view (Char. 241: 1 -> 2).
The genus Sarcosuchus shows the following characters: the posterior dentition of maxillary and dentary present a weak-developed ridges on the enamel surface (Char. 268: 1 -> 0); the maxilla–dentary anterior dentition has a bulbous crown shape (Char. 270: 0 -> 1); the dentary alveoli have a transitional morphology from circular to oval (Char. 272: 0 -> 1); the dentary alveoli one and two are close to each other (Char. 276: 0 -> 1); and the third dentary alveolus is medial in relation to the fourth (Char. 279: 2 -> 0). Clade ( Terminonaris (( Elosuchus + Vectisuchus ) ( Dyrosauridae )) is supported by the following features: the surface below the dental groove is ornamented (Char. 18: 1 -> 0); the rostrum is subequal (Char. 30: 4 -> 2); the frontal surpass the anterior portion of the prefrontals (Char. 96: 1 -> 0); the parietal–postorbital suture is outside the supratemporal fossa (Char. 107: 1 -> 0); the retroarticular process is long (Char. 240: 0 -> 2); the premaxilla–dentary has an aligned occlusion pattern (Char. 246: 1 -> 0); the ulna is shorter than the humerus (Char. 297: 0 -> 1); and the centrum of dorsal vertebrae is spool-shaped (Char. 330: 0 -> 1). The (( Elosuchus + Vectisuchus ) ( Dyrosauridae )) lineage is united by: the posterodorsal process of the premaxilla is anterior to the third maxillary alveolus (Char. 53: 1 -> 0); the ventral margins of maxilla and premaxilla are at the same level (Char. 56: 2 -> 1); there are occlusion pits for first dentary teeth in the premaxilla (Char. 58: 0 -> 1); there is no lacrimal posteroventral process beneath the orbits (Char. 85: 1 -> 0); the upper temporal bar is oblique in relation to the median sagittal axis (Char. 127: 0 -> 1); and the pterygoid composes the posterior, lateral and partially the anterior margins of choana (Char. 208: 1 -> 2). Finally, the ( Elosuchus + Vectisuchus ) clade has the following synapomorphies: the periorbital bones are laterodorsally elevated (Char. 26: 0 -> 1); the orbits are anteriorly inclined (Char. 34: 0 -> 1); the mandibular fenestra is larger than the orbits (Char. 213: 0 -> 1); the crown shape of maxilla–dentary anterior dentition is bulbous (Char. 270: 0 -> 1); and the dorsal osteoderms has a longitudinal keel (Char. 340: 1 -> 0). The features that support Dyrosauridae are available from the Supporting Information, Supplementary Information X.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.