
Tyrannosaurus rex, Osborn, 1905
|
publication ID |
https://doi.org/ 10.5281/zenodo.3898946 |
|
DOI |
https://doi.org/10.5281/zenodo.5204538 |
|
persistent identifier |
https://treatment.plazi.org/id/03E1879F-023C-0C04-E75C-A83CFB48F94A |
|
treatment provided by |
Jeremy |
|
scientific name |
Tyrannosaurus rex |
| status |
|
The primary specimen used in this study was CMNH 7541 , a nearly complete skull of a probably juvenile tyrannosaurid, collected from the Hell Creek Formation of Carter County, Montana (see Gilmore, 1946, for collection details and geological setting). A variety of other tyrannosaurid specimens were studied in connection with this project (for a full listing, see Witmer and Ridgely, 2009). In addition to general observation, the major tool used in this study was CT scanning (computed x-ray tomography), followed by 3D analysis and visualization. CMNH 7541 has been CT scanned no fewer than three times.
The first scans were done in 1990 on a medical scanner at a Toronto hospital, in the presence of Michael Williams, Andrew Leitch, and Robert T. Bakker ( Bakker, 1992). Scanning details have not been published, but results were presented in a popular article by Bakker (1992), who reconstructed the brain cavity and, within the nasal cavity, an elongate turbinate. The same CT data were used later by J. A. Ruben to refute the finding of respiratory turbinates in this skull ( Ruben, 1996; Ruben et al., 1996, 1997).
The Cleveland skull was scanned for the second time in 1998 at the Boeing Rocketdyne CT Lab in California under the auspices of the Cleveland Museum of Natural History with funding from the Field Museum of Natural History. The skull was scanned at a slice thickness of 1 mm with slices in the horizontal plane, yielding 300 slices; voxel sizes were 0.4042 X 0.4042 X 1.0 mm. As part of the collaboration with M. Williams, the Cleveland Museum of Natural History provided Witmer with the full 16-bit dataset from these scans.
In hopes of revealing aspects of anatomy that are unclear in the Boeing dataset, the skull was scanned for the third time in 2005 at O’Bleness Memorial Hospital, Athens, Ohio, on a General Electric LightSpeed Ultra MultiSlice CT scanner equipped with the Extended Hounsfield option, which enhances the ability to resolve details from dense objects such as fossils. Because the Boeing dataset was derived from scans of the skull in the horizontal plane, the x-rays had to pass through the full length of the skull, which can produce artifacts. Thus, we scanned the skull axially, which minimized the amount of material the x-rays had to penetrate. The full skull was scanned helically with a slice thickness of 1.25 mm at 140 kV and 300 mA, yielding 483 slices and voxel sizes of 0.082 X 0.082 X 1.25 mm. The skull was then scanned again, this time focusing on the braincase, tightening the field of view and extending from the caudal end of the skull through the orbits and ending rostrally just in front of the lacrimals’ jugal processes; slice thickness was 625 microns (0.625 mm) at 140 kV and 170 mA, yielding 385 slices and voxel sizes of 0.049 X 0.049 X 0.625 mm. No CT scan is free from artifact, and both the Boeing and Athens datasets have their advantages and disadvantages. Ultimately, much of the work presented here is based on a composite ‘‘superdataset’’ created by registering and combining the Boeing and multiple Athens datasets.
Viewing, analysis, and visualization of the scan data were done using Amira 3.1.1 and 4.2 (Mercury-TGS, Chelmsford, MA) on 32- and 64-bit Windows XP PCs equipped with nVidia Quadro FX graphics cards and 2–8 GB of RAM. Structures of interest were highlighted (segmented) using Amira’s segmentation tools and visualized either in isolation or combined with other structures or the whole skull. Both surfaces and volumes were generated, and these served as the basis for most of the illustrations in this article. Additional information, such as interactive movies and 3D PDFs, are available on the authors’ website: www.ohio.edu/witmerlab.
Museum abbreviations used in this paper are: AMNH, American Museum of Natural History, New York City, New York; BMR, Burpee Museum of Natural History, Rockford, Illinois; CMNH, Cleveland Museum of Natural History, Cleveland, Ohio; FMNH, Field Museum of Natural History, Chicago, Illinois; MOR, Museum of the Rockies, Bozeman, Montana; OMNH, Oklahoma Museum of Natural History, Norman, Oklahoma; ROM, Royal Ontario Museum, Toronto, Ontario; and TMP, Royal Tyrrell Museum of Palaeontology, Drumheller, Alberta.
Results
CMNH 7541 has been the subject of at least three major anatomical studies ( Gilmore, 1946; Bakker et al., 1988; Carr, 1999), and thus there is no need for in-depth anatomical description here. Instead, our intent is to focus on a few areas that have not been adequately discussed in print and that are revealed by the CT scan data. It is also our intent to capitalize on the new-found visualization capabilities provided by the new imaging technologies and software, and so we emphasize illustration over text (in the spirit of the old adage of a picture being worth a thousand words).
General attributes
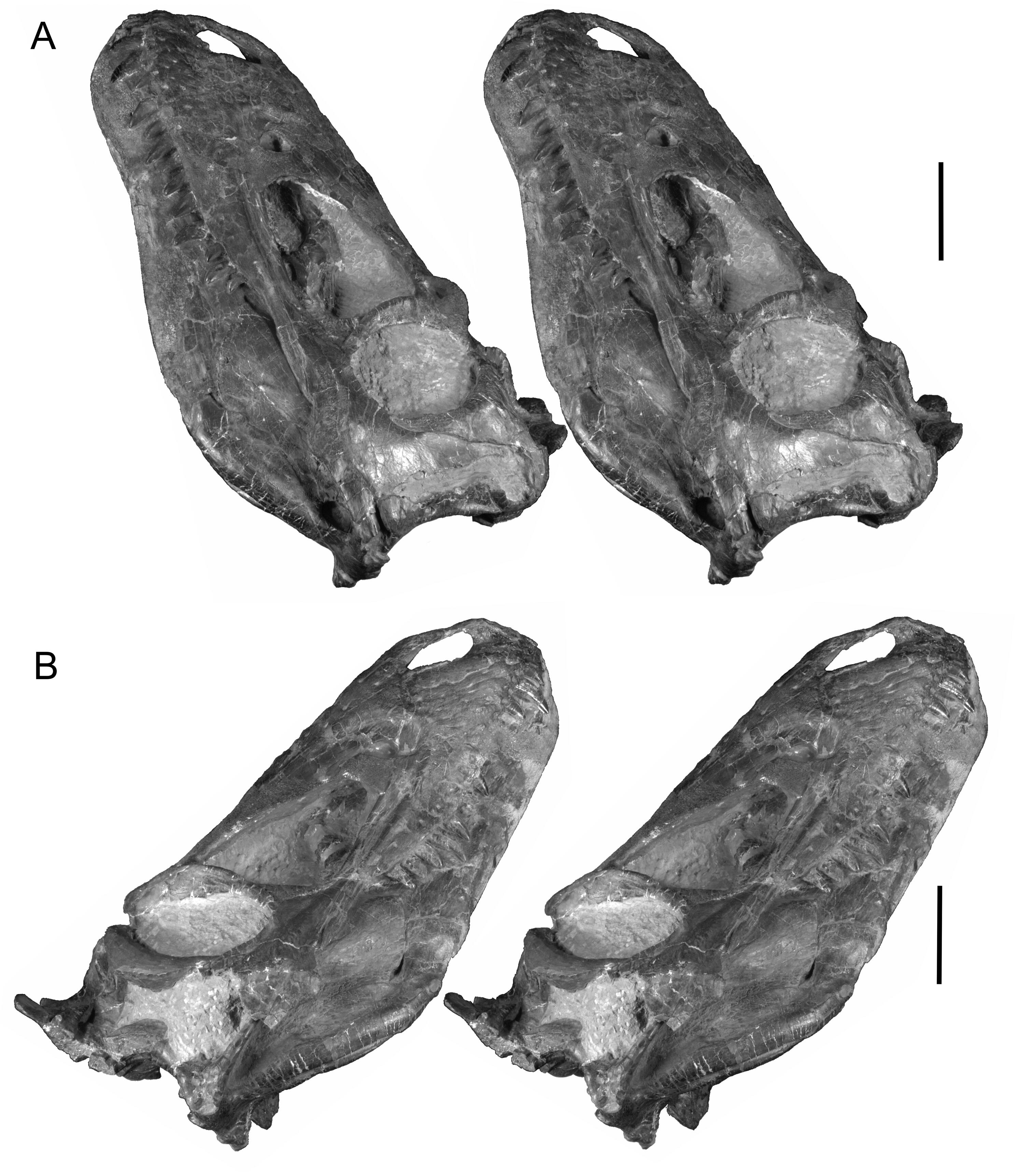
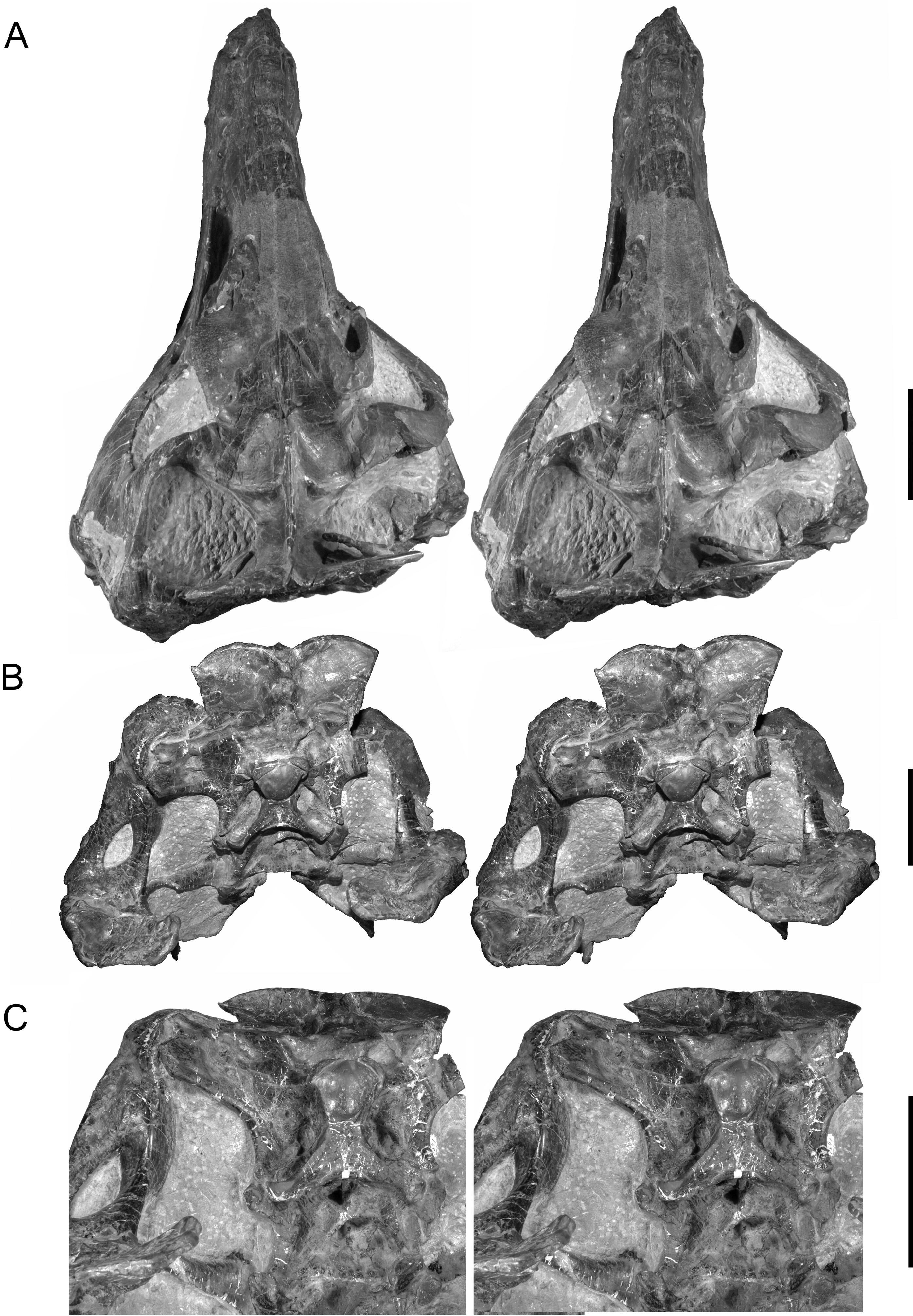
Figures 1–3 View Figure 1 View Figure 2 View Figure 3 provide stereophotographs of the skull in several views, showing not just bony structure but also the amount of matrix remaining in the skull and, to a certain extent, the amount of plaster restoration. Plaster is relatively low density and homogenous and is easily identified and removed in volume renderings of the CT data. Figure 4 View Figure 4 presents views of the skull with the plaster (and some regions of thin matrix) dropped out. Examination of Figure 4 View Figure 4 reveals that the snout had been fractured along an oblique transverse plane running caudodorsally to rostroventrally (arrows in Figure 4C View Figure 4 ) such that, when repaired and restored in plaster (quite expertly, it may be said), much of the central parts of the nasal and antorbital cavities wound up being largely plaster.
Braincase
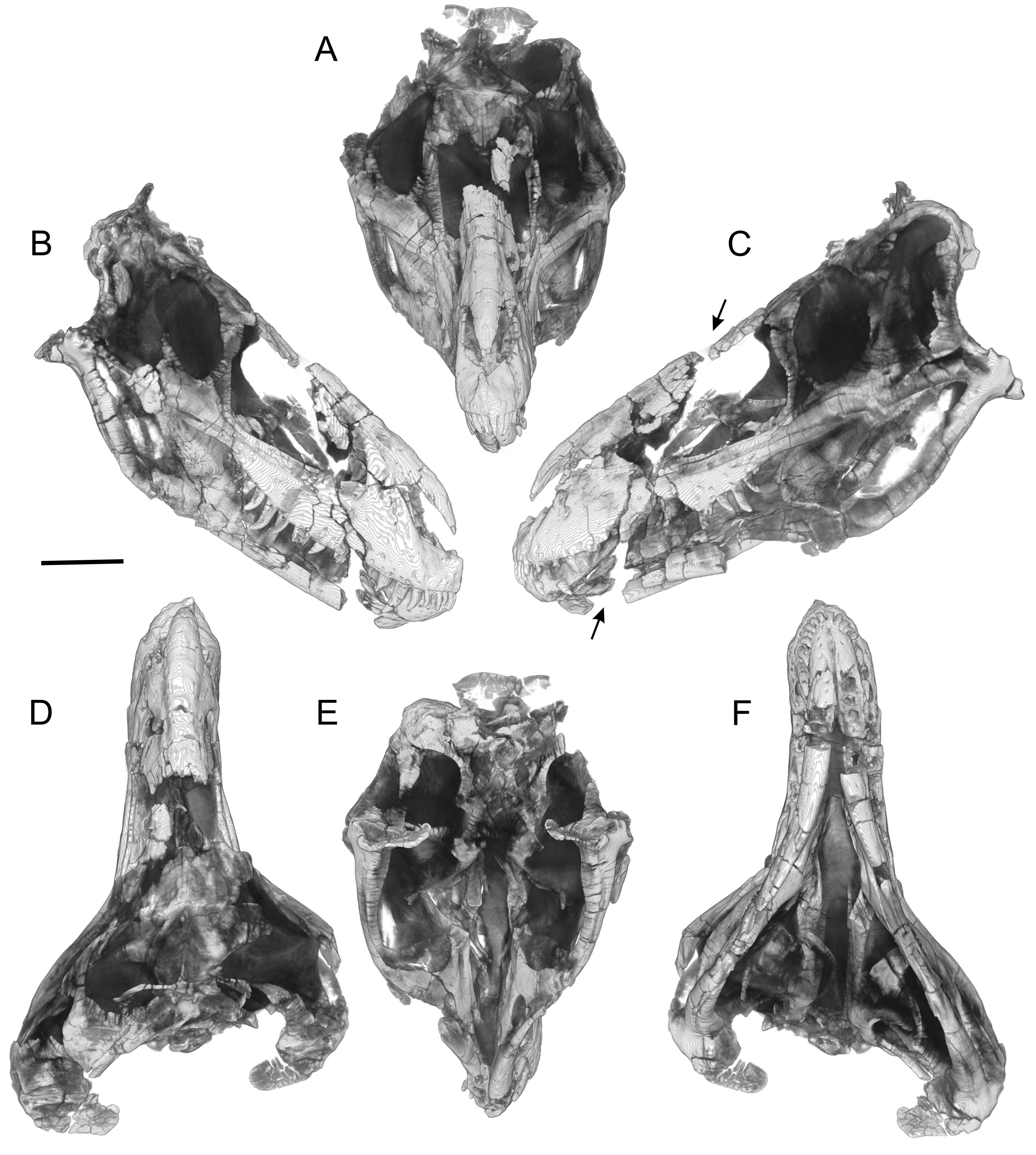
The braincase of CMNH 7541 is visible in dorsal, caudal, and ventral views in the actual (physical) specimen ( Figures 2 View Figure 2 D–E, 3A–C), but the other surfaces are obscured by matrix and other bones. To more easily compare the braincase to those of other theropods, the braincase of CMNH 7541 was digitally ‘‘disarticulated’’ along sutures and then extracted so that it could be viewed in isolation ( Figures 5–8 View Figure 5 View Figure 6 View Figure 7 View Figure 8 ). The general conformation of the braincase is fairly similar to those of other tyrannosaurs in having a well developed otosphenoidal crest (= crista prootica) running from the otoccipital bone caudally and arching rostrally and then ventrally on the prootic, laterosphenoid, and basisphenoid bones ( Figure 6B View Figure 6 ). Tucked ventrally or caudoventrally below the margin of the otosphenoidal crest, from caudal to rostral, are the caudal tympanic recess aperture, columellar recess (with the columella [= stapes] in place in the fenestra vestibuli), the maxillomandibular and facial foramina (for CN V 2–3 and CN VII, respectively), the rostral tympanic recess, and cerebral carotid foramen. As in other tyrannosaurids, the facial foramen opens so close to the maxillomandibular foramen that the two essentially share an aperture laterally ( Witmer et al., 2008; Witmer and Ridgely, 2009; Brusatte et al., 2009).
As in other tyrannosaurids, perhaps all coelurosaurs, and many tetanurans ( Sampson and Witmer, 2007; Witmer and Ridgely, 2009), the ophthalmic nerve (CN V 1) exits the laterosphenoid rostrally via its own foramen. Another attribute shared with other tyrannosaurids is the almost complete ossification of the front of the braincase, with orbitosphenoid and sphenethmoid ossifications contacting their fellows in the midline and enclosing foramina for the olfactory bulbs and tracts, optic nerves (CN II), oculomotor nerves (CN III), and abducens nerves (CN VI) ( Figures 6 View Figure 6 , 8 View Figure 8 ). The sphenethmoid is particularly significant because it preserves details relating to the olfactory apparatus. The sphenethmoid itself is divided by a midline osseous septum (mesethmoid), which in life would have separated the rostral terminations of the olfactory tracts (i.e., the olfactory bulbs), as described for other theropods ( Sampson and Witmer, 2007; Ali et al., 2008; Witmer and Ridgely, 2009). Lateral to the region of the olfactory bulbs are a series of thin bony laminae descending from the roof of the sphenethmoid and possibly also the frontal ( Figures 6A, B View Figure 6 ; 7B, C View Figure 7 ; 8 View Figure 8 ). These laminae are external to the neural domain and would be within the nasal cavity, and thus these laminae are best interpreted as olfactory turbinates that would have supported the sensory olfactory epithelium, as observed in extant taxa. The olfactory apparatus indeed seems well developed in CMNH 7541 , which is consistent with its large olfactory bulbs, as well as with the large bulbs and expansive nasal olfactory regions seen in other tyrannosaurs ( Witmer et al., 2008; Witmer and Ridgely, 2009).
CMNH 7541 shares with other tyrannosauroids modestlysized but widely-spaced basal tubera ( Bakker et al., 1988; Carr, 1999; Currie et al., 2003; Li et al., 2010), yet it retains strong basipterygoid processes. Likewise, all tyrannosaurids, including CMNH 7541 , share extensive pneumaticity associated with the middle ear and pharynx (rostral and caudal tympanic recesses, basisphenoid recesses, subcondylar recess, subsellar recess; Witmer, 1997b; Witmer and Ridgely, 2009; see below).
Despite the similarities with other tyrannosaurids noted above, CMNH 7541 displays a number of noteworthy differences. For example, the cultriform process (parasphenoid rostrum) is relatively low and straight ( Figures 5A View Figure 5 , 6A View Figure 6 ), rather than, as in most other tyrannosaurids ( Russell, 1970; Carr, 1999; Brochu, 2003; Currie, 2003b), having a strongly arched ventral margin that sweeps dorsally before leveling off parallel to the frontals. This may relate to CMNH 7541 having a relatively smaller subsellar recess (a ventral pneumatic chamber in the base of the cultriform process; Figures 6B View Figure 6 , 9 View Figure 9 ) relative to other tyrannosaurids. Among tyrannosaurids, the conformation of the cultriform process and subsellar recess is most similar to that of Gorgosaurus ( Witmer and Ridgely, 2009) and Alioramus ( Brusatte et al., 2009). Another difference is that CMNH 7541 has a much less projecting preotic pendant ( Figure 6A View Figure 6 ), whereas in other tyrannosaurids it is a large and rugose structure associated with the otosphenoidal crest ( Holliday and Witmer, 2008; Witmer and Ridgely, 2009); its weak development in CMNH 7541 may relate to the small size of the animal.
Other differences relate to the location of the vagus foramen (CN X) on the occiput and to the structure of the subcondylar recesses of the basicranium. In most other tyrannosaurids (certainly T. rex ), the vagus foramen (CN X) is located medially in the caudal surface of the otoccipital within a paracondylar pocket or recess adjacent to the occipital condyle. In CMNH 7541 , however, the vagus foramen is located more laterally ( Figure 6C View Figure 6 ), in a position more typical of other coelurosaurs.
Ornithomimids, all tyrannosaurids (although apparently not the tyrannosauroid Xiongguanlong; Li et al., 2010), and potentially even some non-coelurosaurs ( Sampson and Witmer, 2007) have subcondylar recesses (lateral and medial), which are pneumatic recesses located ventral to the occipital condyle and which excavate the basioccipital and otoccipital in the region above the basal tubera. Furthermore, in most tyrannosaurids, there are pneumatic apertures in the floor of the recess leading into chambers within the basioccipital and otoccipital ( Currie, 2003b; Witmer and Ridgely, 2009). The subcondylar recesses of CMNH 7541 differ from those of T. rex . In the former, the recesses as a whole are deeper and the basioccipital and otoccipital pneumatic apertures (leading into the medial and lateral subcondylar sinuses, respectively) are much closer together (essentially adjacent to each other and within a shared fossa; see Figures 5C View Figure 5 , 6C View Figure 6 , 9 View Figure 9 ). Related to this difference is that the ventromedial wall of the subcondylar recess (the condylotuberal crest; Figures 5C View Figure 5 , 6C View Figure 6 , 9 View Figure 9 ) is much stronger in CMNH 7541 than in specimens of T. rex . Taken together, the subcondylar region of CMNH 7541 is more primitive than that of T. rex and is intermediate between the latter and more basal tyrannosaurids (e.g., Gorgosaurus , ROM 1247; Daspletosaurus , FMNH PR308; see Witmer and Ridgely, 2009) and even more basal theropods (e.g., Acrocanthosaurus , OMNH 10146).
Other workers (e.g., Gilmore, 1946; Bakker et al., 1988) have pointed out the divergent nature of the apertures within the basisphenoid pneumatic recess of CMNH 7541 , but 3D visualization of the pneumatic sinuses helps clarify the situation in comparison to other tyrannosaurids. In general, the basisphenoid recess of CMNH 7541 more closely resembles that of Gorgosaurus and Daspletosaurus in having a longer rostrocaudal distance between the intertuberal and interbasipterygoid laminae (= basituberal and basipterygoid webs, respectively, of Bakker et al., 1988; see Figures 5–8 View Figure 5 View Figure 6 View Figure 7 View Figure 8 ), whereas in T. rex these two laminae are quite closely appressed ( Witmer and Ridgely, 2009). However, what makes CMNH 7541 so divergent is the pattern of apertures within the basisphenoid recess. As preserved ( Figures 3C View Figure 3 , 6C View Figure 6 ), there are three asymmetrical apertures, unlike any other known theropod. When traced dorsally into the pneumatic sinuses using the CT data, the caudal two apertures can be seen to represent a pair (even if they are not fully bilaterally symmetrical) because they expand into paired (i.e., left and right) sinuses (the caudal basisphenoid sinuses; Figures 8C View Figure 8 , 9 View Figure 9 ) that lead into the basioccipital and communicate on either side of the occiput with the medial subcondylar recesses. Expansion of the basisphenoid recess into sinuses within the basioccipital is fairly common in theropods generally, although the pattern of apertures in CMNH 7541 is unique.
The third and most rostral aperture within the basisphenoid recess has been difficult to interpret. Other tyrannosaurids have a rostral pair of foramina (left and right) in the interbasipterygoid lamina that lead into pneumatic chambers within the basisphenoid ( Russell, 1970; Bakker et al., 1988; Witmer, 1997b; Currie, 2003b; Witmer and Ridgely, 2009). Tracing the seemingly single aperture in CMNH 7541 in the CT data reveals that the aperture branches dorsally into essentially left and right sinuses (the rostral basisphenoid sinuses) that expand within the basisphenoid and eventually communicate with the rostral tympanic recesses on their respective sides ( Figures 7C View Figure 7 , 8C View Figure 8 , 9 View Figure 9 ). As a result, it would seem that the pair of foramina in the interbasipterygoid lamina of other tyrannosaurids is present in CMNH 7541 , but that the two apertures share a common fossa that is diverted towards the right side somewhat. Thus in sum, we are proposing that the basisphenoid pneumatic sinuses are fundamentally similar to those of other tyrannosaurids, albeit highly modified. Gilmore (1946, p. 10) explained this unusual and asymmetric morphology as representing ‘‘an unhealthy condition of the bone,’’ but there is no overt sign of pathology in this region.
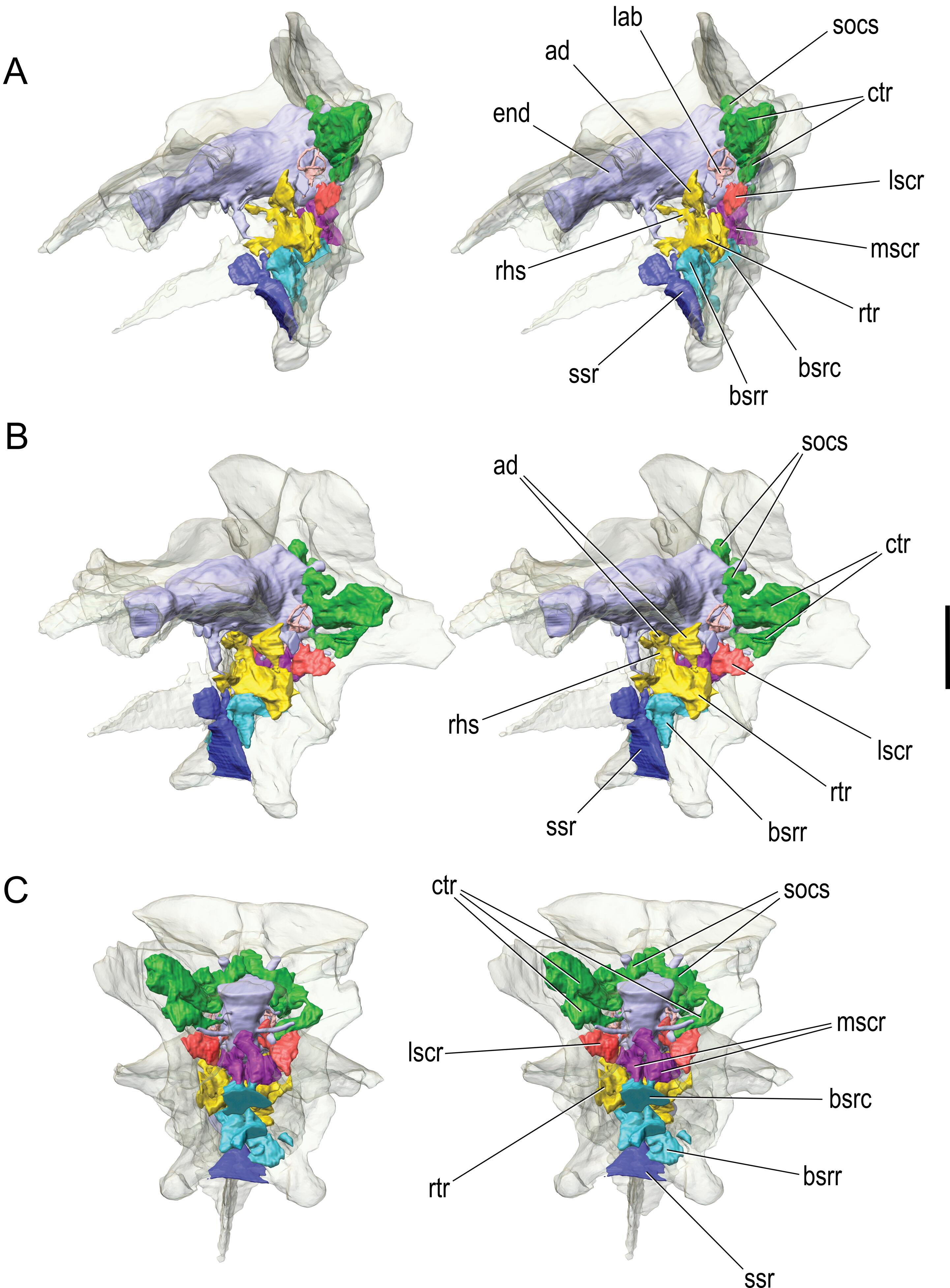
Pneumatic sinuses of the braincase
The pneumatic recesses of CMNH 7541 have been touched on above, but they will receive fuller treatment here in that they have been reconstructed in three dimensions using the CT-scan data ( Figure 9 View Figure 9 ). As noted, there is a clear aperture leading into the caudal tympanic recess, and, as in most other coelurosaurs ( Witmer, 1997b), the aperture is located in the base of the paroccipital process adjacent to the columellar recess and bounded by the prootic and opisthotic. In CMNH 7541 , the caudal tympanic recess expands within the paroccipital process, giving off two diverticula, one ventrally and the other dorsally. The ventral diverticulum enters the crista tuberalis (= basioccipital wing of the otoccipital, descending ventral root of the paroccipital process of Bakker et al., 1988) where it is broadly confluent with the lateral subcondylar recess. Despite this confluence, the cavity within the crista tuberalis of CMNH 7541 clearly derives as a primary diverticulum of the lateral subcondylar recess, because, in other tyrannosaurids (e.g., T. rex , Gorgosaurus ), the communication between the two pneumatic spaces is slight to nonexistent, and it is the lateral subcondylar sinus that occupies the crista tuberalis ( Witmer and Ridgely, 2009). The dorsal diverticulum certainly derives as an outgrowth of the caudal tympanic recess and forms a series of interconnecting chambers medially within the otoccipital and supraoccipital and laterally within the paroccipital process ( Figure 9 View Figure 9 ), as in other tyrannosaurids and indeed most other coelurosaurs ( Witmer, 1997b; Witmer and Ridgely, 2009). There may be a tenuous communication between contralateral recesses within the supraoccipital but not the broad communication observed in T. rex and Gorgosaurus .
The rostral tympanic recess is located, again fairly typically, in the region where the cerebral carotid artery enters the braincase under shelter of the otosphenoidal crest, that is, in the area where the basisphenoid, prootic, and laterosphenoid contact each other ( Figure 9 View Figure 9 ). The rostral tympanic recess expands within the basisphenoid where it communicates with both the rostral and caudal basisphenoid sinuses, but more broadly with the rostral basisphenoid sinus. The rostral tympanic recess has a major dorsal diverticulum (the ascending diverticulum; Figure 9A, B View Figure 9 ) that leads into the substance of the laterosphenoid bone, occupying the region between the canals for the ophthalmic nerve (CN V 1) rostrally and maxillomandibular nerve (CN V 2–3) caudally. The connection between the ascending diverticulum and the rostral tympanic recess is very narrow. The ascending diverticulum is present in Gorgosaurus but is usually absent (or very rudimentary) in adult T. rex and Dapletosaurus ( Witmer and Ridgely, 2009), which could be a legitimate, systematically informative difference. However, a newly discovered, very young skull of Tarbosaurus (just 29 cm total skull length; Tsuihiji et al., 2007, in review) displays a well developed ascending diverticulum of the rostral tympanic recess; although we lack comparable CT data for adult Tarbosaurus , if we assume that Tarbosaurus resembled adult T. rex , ontogenetic loss of the ascending diverticulum in tyrannosaurines remains a possibility.
The rostral tympanic recess has another, more medial, diverticulum (the retrohypophyseal sinus, Figure 9A, B View Figure 9 ) that extends dorsomedially within the clivus of the basisphenoid just caudal to the hypophyseal (pituitary) fossa. This sinus represents a contralateral communication of the left and right rostral tympanic recesses, and has been found in all the tyrannosaurids studied here ( Witmer and Ridgely, 2009). In CMNH 7541 , the retrohypophyseal sinus itself sends a small median diverticulum between the paired abducens (CN VI) canals.
The sinuses associated with the basisphenoid recess have been mentioned above in connection with their external apertures. Given the asymmetry of their bony apertures, it is not surprising that the sinuses themselves are quite asymmetrical, much more so than the caudal and rostral tympanic recesses and their associated diverticula. The caudal basisphenoid sinuses expand dorsally and somewhat caudally, passing through the basioccipital, as noted above, to become broadly confluent with the medial subcondylar recess on the occiput ( Figure 9 View Figure 9 ). The caudal basisphenoid sinus has some communication with the rostral tympanic recess (more so on the left side). The rostral basisphenoid sinuses expand dorsally and rostrally within the basisphenoid. Interestingly, although the shared aperture of the two rostral basisphenoid sinuses within the recess is shifted to the right side, the left sinus is larger and more broadly communicates with the rostral tympanic recess ( Figure 9 View Figure 9 ). This asymmetry could be natural but some small amount of postmortem crushing cannot be ruled out, as the right side of the braincase is generally not as well preserved as is the left side.
The subsellar recess is located rostral to the interbasipterygoid lamina at the ventral base of the cultriform process ( Figure 9 View Figure 9 ). The subsellar recess is relatively small in comparison to that of T. rex and Daspletosaurus but is similar in size to that of some juvenile Gorgosaurus specimens (e.g., ROM 1247). There are no pneumatic foramina within the subsellar recess on CMNH 7541 .
On the right side of CMNH 7541 , but not the left, there is an aperture in the basisphenoid bone just dorsal to the base of the basipterygoid process. The aperture is surrounded by a shallow fossa, and these features can be regarded as a basipterygoid recess ( Chure and Madsen, 1996; Witmer, 1997b). The aperture leads into a sinus (the basipterygoid sinus) that ascends dorsally within the basisphenoid. Its communications with other sinuses in the basisphenoid are slight to nonexistent, although it does appear to breach the interbasipterygoid lamina within the basisphenoid recess, where, if so, it would then communicate with the air sinus located there. The basipterygoid process itself is not pneumatized.
Finally, the subcondylar recesses open into a pneumatic fossa on the occiput below the occipital condyle ( Witmer, 1997b) and have been mentioned above in connection with pneumatic apertures located in the otoccipital and basioccipital within the fossa ( Figure 9C View Figure 9 ). Carr (1999) and Currie (2003b) both regarded the fossa for the subcondylar recesses of CMNH 7541 as shallow, but to our eyes it seems intermediate between the deep recess of young Gorgosaurus specimens (ROM 1247) and the very shallow recess of mature specimens of T. rex (e.g., AMNH 5027, AMNH 5117, FMNH PR2081; see Witmer and Ridgely, 2009). As noted above, the otoccipital aperture of the lateral subcondylar recess expands into a sinus within the crista tuberalis (where it communicates with the caudal tympanic recess), whereas the basioccipital aperture leads to the medial subcondylar sinus that communicates with the caudal basisphenoid sinus. The medial subcondylar sinus does not pneumatize the occipital condyle but just the very base of the neck, as Osborn (1912) showed for T. rex (AMNH 5029; see also Witmer and Ridgely, 2009).
Thus in summary, CMNH 7541 has an extensively pneumatized braincase, with clearly identifiable rostral and caudal tympanic recesses, a series of basisphenoid sinuses arising rostrally and caudally from the basisphenoid recess, medial and lateral subcondylar recesses, a small subsellar recess, and, on one side only, a basipterygoid recess. Virtually all of these sinuses communicate broadly with adjacent sinuses. The two tympanic recesses are clearly derived from the middle ear sac (as their names imply), but the basisphenoid and subsellar recesses may derive from a separate median pharyngeal system ( Witmer, 1997b). Likewise, although the subcondylar recesses could be tympanic in origin, it is not possible to rule out pneumatization via a cervical pulmonary diverticulum ( Witmer, 1997b; Witmer and Ridgely, 2009).
Columella (= stapes)
The columella is clearly preserved in natural position on the left side ( Figure 6A, B View Figure 6 ; 8A, B View Figure 8 ). It is a very delicate element, extending from the fenestra vestibuli within the columellar recess, passing along the paroccipital process below the otosphenoidal crest, to end laterally just caudomedial to the dorsal head of the quadrate. The columella is only about 1.4 mm in diameter, yet is 71.7 mm in length. The length is probably an underestimate in that the lateral tip is exposed in the actual fossil, and it is unknown how much was lost in collection, preparation, etc. The fact that the columella is preserved in situ and is straight suggests that the braincase region as a whole has not undergone significant deformation.
Quadrate and its pneumaticity
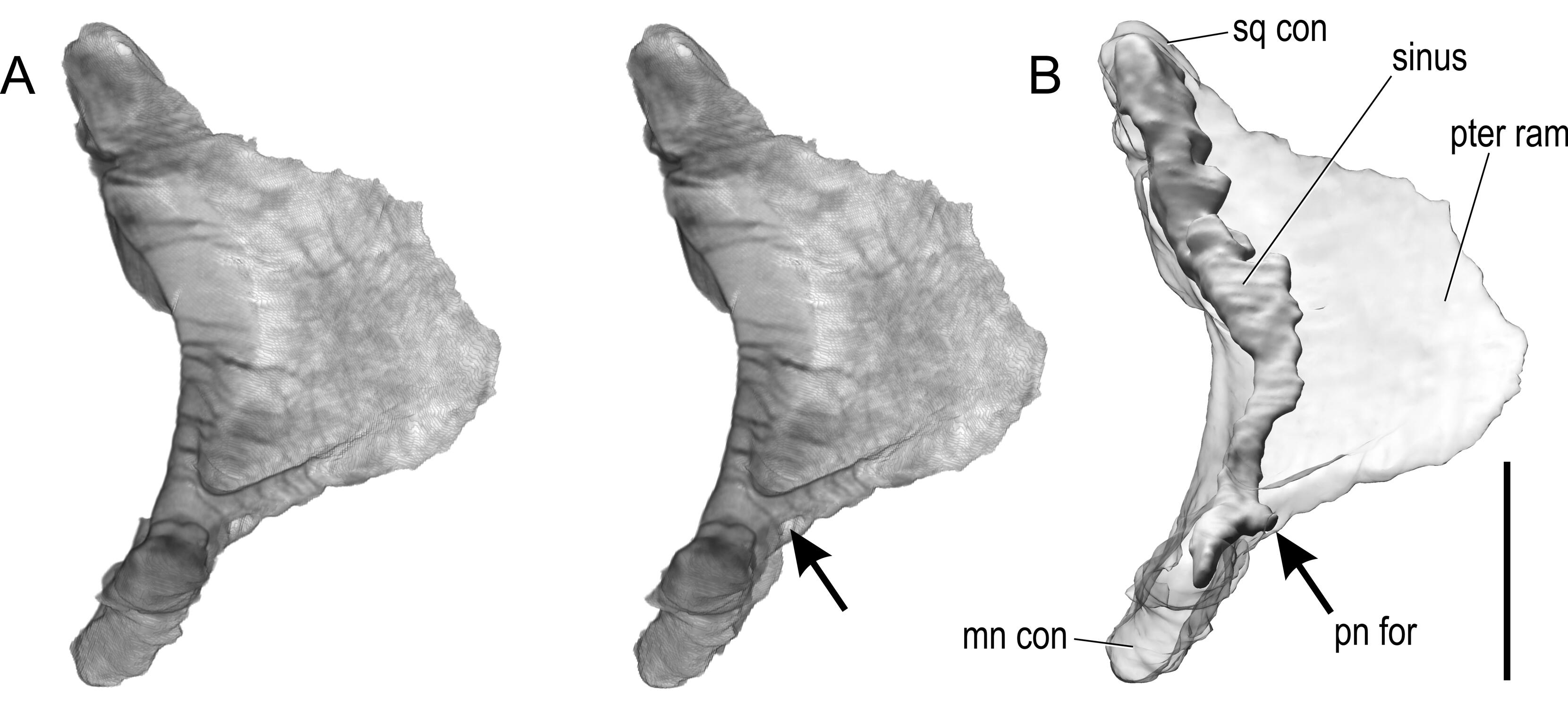
The left quadrate was digitally extracted from the full CT dataset to examine and illustrate the position of its pneumatic aperture ( Figure 10 View Figure 10 ). Although a taxonomic difference in position of this foramen has not been noted previously, the quadrate pneumatic foramen of albertosaurines ( Gorgosaurus : AMNH 5363; Albertosaurus : TMP 81.10.1; see also Currie, 2003b, p. 200) is in a slightly different position than in Daspletosaurus (FMNH PR 308) and Tyrannosaurus (FMNH PR2081). In the former, the pneumatic foramen is directed more medially, whereas in the latter group the crest of bone above the foramen on the quadrate’s pterygoid ramus is much stronger and diverts the pneumatic foramen to a more rostral position such that it no longer directly faces the tympanic cavity. In CMNH 7541 , the quadrate pneumatic foramen again has more of an intermediate position ( Figure 10 View Figure 10 ). The foramen itself is relatively small in CMNH 7541 . BMR P2002.4.1 is very similar in these regards. The quadrate pneumatic sinus in CMNH 7541 is somewhat better preserved on the right side (not illustrated in Figure 10 View Figure 10 ), but on both sides the sinus extends essentially the full height of the element ( Figure 10B View Figure 10 ).
Quadratojugal and its pneumaticity
The quadratojugal is preserved on only the left side of CMNH 7541 , and even here the bone is fractured and displaced such that its jugal process remains in articulation with the jugal, whereas its squamosal process has been shifted dorsally and rostrally. The most significant attribute of the quadratojugal is the peculiar foramen in its lateral surface within the ventral apex of the lateral fossa. BMR P2002.4.1 shows a virtually identical foramen, but such a foramen is absent in adult T. rex and has not been reported in other tyrannosaurids, other than a small foramen in a specimen of Gorgosaurus sp. reported by P. Larson (2008). Although only rarely noted in print (e.g., P. Larson, 2008), some tyrannosaur workers have wondered if this difference supports the validity of the taxon Nanotyrannus lancensis . To clarify the anatomy, we examined this foramen in the CT dataset and digitally extracted the bone for visualization ( Figure 11 View Figure 11 ). The foramen is almost certainly a pneumatic foramen, expanding into a blind cavity within the substance of the bone. The cavity has no medial outlets, only the lateral aperture. This also would tend to indicate that the lateral quadratojugal fossa is a pneumatic fossa, most likely associated with the middle ear. Although it remains possible that presence of this quadratojugal pneumatic foramen is a juvenile feature of T. rex that was lost later in ontogeny, the very young skull of Tarbosaurus described by Tsuihiji et al. (2007; in review) lacks such a foramen, as do adult Tarbosaurus quadratojugals, suggesting that presence and then loss of the pneumatic foramen was not a general ontogenetic sequence of tyrannosaurines.
Palatine bone and its pneumaticity
The left palatine bone of CMNH 7541 also was digitally extracted ( Figure 12 View Figure 12 ). The bone is a fairly typical tyrannosaurid palatine. The bone is pneumatic as in other tyrannosaurids and some other theropods ( Witmer, 1997a, b; Witmer and Ridgely, 2008b) with two pneumatic foramina plainly visible on the actual skull ( Figure 2A View Figure 2 ). When examined with CT, however, it can be seen that the two pneumatic sinuses within the bone do not communicate but instead form separate chambers. Although the vomeropterygoid process is fully pneumatized ( Figure 12B View Figure 12 ), the bone is not strongly inflated, as seen in many T. rex specimens (e.g., FMNH PR2081, MOR 008). In general, the palatine of BMR P2002.4.1 has a similar conformation with a large pneumatic aperture caudally, but, unlike CMNH 7541 , the rostral pneumatic feature does not breach the bone and so is a fossa, not a foramen.
Vomer
The shape of the rostral end of the vomer is an important character in tyrannosaurid systematics, with basal tyrannosauroids and albertosaurines ( Gorgosaurus and Albertosaurus ) retaining the primitive condition of a narrow, ‘‘lanceolate’’ vomer and tyrannosaurines ( Daspletosaurus , Tarbosaurus , Tyrannosaurus ) having a derived, laterally expanded, diamond-shaped vomer ( Holtz, 2001, 2004; Currie et al., 2003; Li et al., 2010). The vomer of CMNH 7541 is almost completely enclosed in matrix, with just the caudalmost portion exposed within the antorbital cavity ( Figures 1A View Figure 1 , 2A View Figure 2 ). So, to shed light on the systematically important end of the bone, we digitally extracted the vomer for visualization ( Figure 13 View Figure 13 ). The vomer of CMNH 7541 has the primitive condition, with a narrow rostral end. In fact, the rostralmost tip has the further primitive attribute of forking to contact the palatal processes of the maxilla and premaxilla. Overall, the vomer is a fairly typical non-tyrannosaurine theropod vomer with a dorsal sulcus and a ventral keel. The vomer of BMR P2002.4.1 is again almost identical to that of CMNH 7541 . A valid question is whether this morphology is truly primitive or just reflective of an early ontogenetic stage that would transform later in life to the definitive adult tyrannosaurine condition (i.e., T. rex ). In this context, the very young Tarbosaurus specimen described by Tsuihiji et al. (2007; in review) is significant in having a rostrally narrow vomer similar to those of CMNH 7541 and BMR P2002.4.1 , suggesting that indeed a lanceolate vomer can ontogenetically transform into a diamond-shaped vomer. That said, if CMNH 7541 pertains to a juvenile T. rex , then the rate of transformation would have had to have been markedly different in T. rex and Tarbosaurus in that an unnumbered juvenile Tarbosaurus skull (only about 27% larger than CMNH 7541 ) already has a diamond-shaped vomer.
Possible respiratory turbinates
Respiratory turbinates in amniotes have received a great deal of attention because of their potential role in debates on dinosaur metabolic status ( Bakker, 1992; Ruben, 1996; Ruben et al., 1996, 1997). The idea promoted by J. A. Ruben (1995) is that respiratory turbinates may be mandatory in endothermic animals to help mitigate the effects of respiratory evaporative water loss that would result from the high lung-ventilation rates characteristic of endotherms. As mentioned above, CT scanning of CMNH 7541 has been a major player in this debate. Bakker (1992) reconstructed a large scrolled turbinate within the snout of this animal, but, given that the reconstruction was published in a popular magazine (Discover), no details were given. Later Ruben and his team (1996, 1997) argued (using the same scan data) that no turbinates were present, and that, moreover, the airway was too small to house turbinates.
Unfortunately, CMNH 7541 is not the best specimen on which to base a debate, because so much of the snout is damaged and reconstructed with plaster. As noted above, clear olfactory turbinates are present far caudally ( Figures 5–8 View Figure 5 View Figure 6 View Figure 7 View Figure 8 ); they are immediately adjacent to the olfactory bulb fossa and could not have functioned as respiratory turbinates because they are well out of the main nasal airstream (see Witmer and Ridgely, 2008b, 2009). Rostral to the olfactory turbinates is a large segment of matrix in which no ossified or calcified (i.e., fossilized) turbinatelike structures can be seen, and rostral to this is a large plasterreconstructed area (in association with the oblique transverse fracture mentioned earlier; see Figure 4B, C View Figure 4 ). Ironically, this large region is the area in which Bakker (1992, p. 61) reconstructed his turbinates.
Our new scanning of CMNH 7541 , however, has identified structures within the rostral portion of the snout—in front of the region where Bakker (1992) reconstructed a turbinate and behind the region where Ruben et al. (1996) showed turbinates to be lacking—that may be interpreted as being respiratory turbinates ( Figure 14 View Figure 14 ). The structures in question are located in the main nasal airway above the maxillary antra (which themselves are largely reconstructed in plaster). These structures form a series of thin, but moderately dense laminae visible within undistorted matrix that are associated with both the nasal and the maxilla and project into the nasal airway. There are some regions of symmetry, and some elements that can even be tentatively regarded as ‘‘scrolls’’ ( Figure 14 View Figure 14 ). The density of these putative turbinates is less than the adjacent nasal and maxilla but more than the surrounding matrix. Whether or not these moderate density values reflect calcification of cartilaginous structures or just very thin bone is difficult to determine. Certainly, they are positioned appropriately to be turbinates with a respiratory function in that they are within the portion of the main nasal airway between the naris and choana.
Mandible
The two mandibles are preserved in full occlusion with the skull, and so, although many details of the mandible can be seen ( Figures 1–3 View Figure 1 View Figure 2 View Figure 3 ), details of the tooth-bearing portions are not easily observed. Thus, obtaining information on the common comparative metric of number of tooth positions had been impossible for CMNH 7541 . To remedy this situation, we digitally extracted the right mandible because the dental region is better preserved ( Figure 15 View Figure 15 ). CMNH 7541 has at least 16 relatively clear tooth positions, which is similar to BMR P2002.4.1 , which has 16 or 17 (P. Larson, 2008). For comparison, Holtz (2004, p. 119) provided a range among tyrannosaurids of 18 in Alectrosaurus and 11 in T. rex . Hurum and Sabath (2003, p. 187) reported 12–14 dentary tooth positions in T. rex and 14–15 in Tarbosaurus . Russell (1970) reported 15–16 for Daspletosaurus . Thus, CMNH 7541 is fully within the range of Tyrannosauridae , but does not overlap T. rex . Significantly, Tsuihiji et al. (2007; in review) reported the exact same range of dentary tooth positions (14–15) in a very young specimen of Tarbosaurus as Hurum and Sabath (2003) reported for adult Tarbosaurus , suggesting no ontogenetic change in tooth counts in this species that is so closely related to T. rex , and thus diminishing the case for CMNH 7541 being a young T. rex .
Discussion
The preceding sections have sought to provide anatomical details of the Cleveland tyrannosaur skull ( CMNH 7541 ) that have not been previously widely available (or available at all). CT scanning—and, more significantly, 3D visualization of the CT scan data—has yielded a new look at this famous fossil. For many workers, the systematic question is paramount, but it is obvious that the ontogenetic question is inextricably linked to any systematic outcome. That is, are the differences observed in CMNH 7541 attributable to its apparent young age or to it being a different taxon or both? Some of the different ideas were presented in the Introduction, and no firm resolution is offered here, in part because other data pertinent to this debate have yet to be published. Most significant is the discovery of a skull and associated partial postcranium ( BMR P2002.4.1 ) of an animal that is very similar to CMNH 7541 and which will be critical in providing information from the postcranial skeleton that could be relevant to the systematic question. Moreover, there are other, still unpublished skulls of young and juvenile tyrannosaurids ( Tsuihiji et al., 2007; in review; N. Larson, 2008; Carr and Williamson, 2010) that can shed light on patterns of ontogenetic transformation and help with discrimination of primitive versus juvenile characters. Other unpublished, clearly adult specimens have raised the prospect of there being species of tyrannosaurids in the Hell Creek other than T. rex (N. Larson, 2008; P. Larson, 2008). The question in this case would be, do these specimens display attributes of CMNH 7541 that might indicate that they are adult Nanotyrannus lancensis ? Until these new specimens are analyzed and published, the status of CMNH 7541 must remain uncertain.
Having said that, the foregoing discussions have revealed an interesting suite of apparently derived attributes observed in CMNH 7541 that might suggest that it represents a taxon separate from T. rex . For example, the patterns of pneumatic sinuses and their bony apertures in the braincase are quite distinct from other tyrannosaurids. Although pneumatic sinuses certainly can change ontogenetically ( Witmer, 1990, 1997a, b; Witmer and Ridgely, 2008b; and references therein), most of the changes in later ontogeny of most taxa pertain to expansion, contraction, and/or communication of sinuses and not to the basic pattern of bony apertures. For example, Gorgosaurus and Tyrannosaurus are more similar to each other in their patterns of basicranial pneumatic foramina than either is to the highly asymmetrical apertures of CMNH 7541 , and there is no evidence to suggest that either of the two named taxa went through an ontogenetic stage resembling CMNH 7541 . Likewise, the broad communication of the caudal tympanic recess and lateral subcondylar recess in CMNH 7541 is unique among tyrannosaurs sampled thus far. To these apparent apomorphies can be added the findings from the brain endocast of CMNH 7541 ( Witmer and Ridgely, 2009), such as the rostrally offset pituitary fossa and orbital cranial nerve canals, as well as the strongly downturned endocast and skull as a whole, as indicated by the orientation of the lateral semicircular canal of the inner ear.
Although many workers have been struck by the T. rex -
like temporal expansion of CMNH 7541 , this derived character is coupled with a series of primitive attributes, such as the narrow and lanceolate vomer, the uncompressed basisphenoid recess, the pattern of pneumatic apertures in the subcondylar recess, the strength of the condylotuberal crest, the low cultriform process and small subsellar recess, the laterally positioned vagus foramen, and the high number of dentary tooth positions. Although the possibility cannot be ruled out, it seems hard to believe that the animal pertaining to CMNH 7541 would have ontogenetically transformed all of these attributes (both primitive and derived) and grown up to be a typical member of Tyrannosaurus rex . CMNH 7541 does not pertain to a hatchling, but rather a fairly mature (‘‘teenage’’) animal, though perhaps not yet in the exponential stage of growth (in the sense of Erickson et al., 2004), and thus it is hard to envision such major morphological changes. Certainly, significant morphological changes can take place in animals at nearly adult age, as Sampson (1999, 2001; see also Sampson et al., 1997) showed for ceratopsids in which many of the attributes relating to sexual display develop at essentially adult body sizes (see also Horner and Goodwin, 2006, 2009). Obviously, few if any of the features listed above for CMNH 7541 are of the type that would either be recruited for sexual display or are even remotely related to secondary sexual characteristics. We ( Witmer and Ridgely, 2009) previously evaluated in some detail other means of explaining our findings, such as diagenetic distortion or pathology, but neither explanation is compelling. That said, we recognize that some aspects of CMNH 7541 are unusual enough that its being simply aberrant may always remain possible, which is why we have never stated definitively that it is not a juvenile T. rex . Ultimately, until the known but unpublished specimens are fully analyzed, we regard the taxonomic status of CMNH 7541 to be an open question, being either a separate taxon ( Nanotyrannus lancensis ) or a juvenile Tyrannosaurus rex .
Finally, given the finding here of structures interpretable as respiratory turbinates, it is tempting to make broader claims about metabolic physiology and endothermy. We hesitate to do so, because the skull has only a small area well enough preserved to make such assessments, and we would prefer to be able to sample more widely within the nasal cavity. Nevertheless, taking our data at face value, it is difficult to explain the laminar structures within the nasal airway as anything else. They are not artifacts of the CT scanning process, nor are they sedimentary structures. In fact, such structures are found nowhere else in the entire dataset. Assuming that they are indeed real structures projecting into the nasal airway and covered with moist mucosal respiratory epithelium, it is hard to imagine how they would not function as counter-current heat exchangers, precisely as postulated by Ruben and colleagues (1996, 1997). Whether or not these structures are indeed causally associated osteological correlates of endothermy or whether these supposed turbinates are extensive enough to have the effect posited by Ruben et al. (1996) is harder to say, but certainly it now seems unreasonable to argue that such structures are absent in dinosaurs.










No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.