
Tipula, Linnaeus, 1758
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4837.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:001B5DF8-A24D-4353-8BEA-8B181BE9D279 |
|
DOI |
https://doi.org/10.5281/zenodo.4454903 |
|
persistent identifier |
https://treatment.plazi.org/id/03E187A2-9322-A87E-6FEB-FC7227B3C636 |
|
treatment provided by |
Plazi |
|
scientific name |
Tipula |
| status |
|
Tipula (Vestiplex Bezzi)
Tipula , subg. Vestiplex Bezzi, 1924: 230 . Type species: Tipula cisalpina Riedel, 1913 (original designation). Additional references: Edwards, 1931: 79; Alexander, 1934b: 396; 1935: 117; 1965: 355; Mannheims, 1953: 116; Savchenko, 1964: 132.
Vestiplex was first proposed by Bezzi (1924) as a subgenus of genus Tipula for the type species T. cisalpin recorded from West Palaearctic ( Italy, Switzerland). Additional characters of T. ( Vestiplex ) were provided by several subsequent authors ( Edwards 1931; Alexander 1934b, 1935; Savchenko 1964).
Females belonging to the subgenus T. ( Vestiplex ) are characterized by the ovipositor with powerful, heavily sclerotised and horizontal cercus, the outer (ventral) margin of which is usually serrate, but smooth in several Asiatic species. The hypovalva is small to rudimentary ( Alexander 1935, 1965; Alexander & Byers 1981). The male genitalia is extremely polymorphic ( Savchenko 1964), typically with tergite 9 forming a shallowly concave and sclerotised saucer, while other species have tergite 9 completely divided longitudinally by a pale membrane ( Alexander 1935; Alexander & Byers 1981).
The subgenus T. ( Vestiplex ) includes 181 extant species which are distributed throughout the Nearctic, Palaearctic and Oriental Regions ( Oosterbroek 2019). No fossil species of T. ( Vestiplex ) are described so far and only Matthews & Telka (1997) mentioned ovipositors of possibly T. ( Vestiplex ) females from the Cape Deceit Formation in Western Alaska, 1.8 MYA.
The species of T. ( Vestiplex ) are separated into various species groups based on morphological structures of the male hypopygium, mainly details of tergite 9. The taxonomy of species with a saucer-shaped tergite 9 is comparatively well investigated by different authors ( Hemmingsen 1956; Mannheims 1953; Theowald & Mannheims 1963; Savchenko 1960, 1964), and seven species groups are recognized: coquillettiana, erectiloba, excisa , leucoprocta , nubeculosa , scripta , and virgatula ( Starkevich 2012) .
Species with divided tergite 9 currently are separated into additional seven species groups ( Starkevich 2012; Starkevich et al. 2019). Edwards (1928) proposed a himalayensis species subgroup of the arctica group for T. (V.) avicularia Edwards, 1928 . Alexander later distinguished a himalayensis species group and included a number of species with divided tergite 9 ( Alexander 1932, 1933, 1934 c, 1935, 1936 b, 1959, 1963). Savchenko (1964) also proposed several species complexes, avicularia , divisotergata, and subtincta, for various Asiatic species based on features of the male hypopygium. Additional species groups proposed based on revisionary study of the subgenus include: bicornigera, deserrata and eurydice ( Starkevich 2012; Starkevich et al. 2019).
Characteristics of Mongolian T. ( Vestiplex ) crane flies
Diagnosis. Mongolian T. ( Vestiplex ) crane flies are medium to large sized, body length of males 11.0–22.0 mm, of females 15.9–31.0 mm. Body coloration varies from yellow to blackish brown ( Figs 1–3 View FIGURES 1–3 ). Antennae (males) length ranges from short, reaching pronotum to elongate, reaching the base of the abdomen if bent backwards. The shape of the flagellomeres varies from simple cylindrical to large basal enlargement and deep incision at middle.
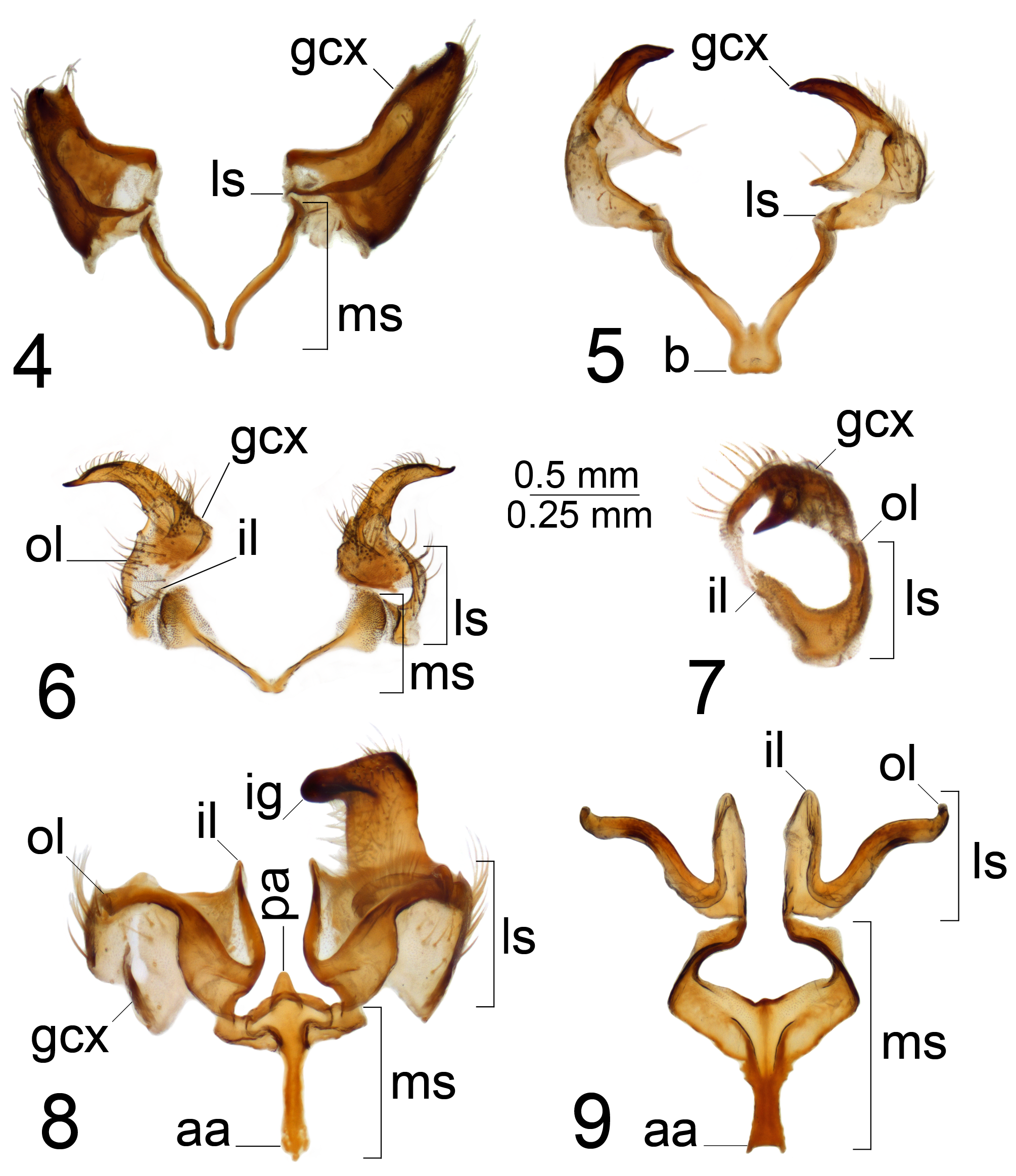
Tergite 9 of the male hypopygium forms a flattish or shallowly concave plate with caudal portion forming a more or less distinct area with narrow rised rim termed the “tergal saucer” ( Alexander 1934b), which varies in shape among the different species groups. The gonocoxite is separated from the sternite by a distinct suture, triangular or rectangular in shape ( T. (V.) virgatula and T. (V.) longitudinalis Nielsen, 1929 ), with an extended and obtuse apex ( T. (V.) nubeculosa Meigen, 1804 ), terminating with a horn or spine ( excisa and scripta groups) or simple without extension ( leucoprocta group). The inner gonostylus is generally a transversely curved, claw-shaped plate with preapical notch, bifid apex or middorsally toothed. The aedeagal guide is tube- or depressed, plate-shaped.
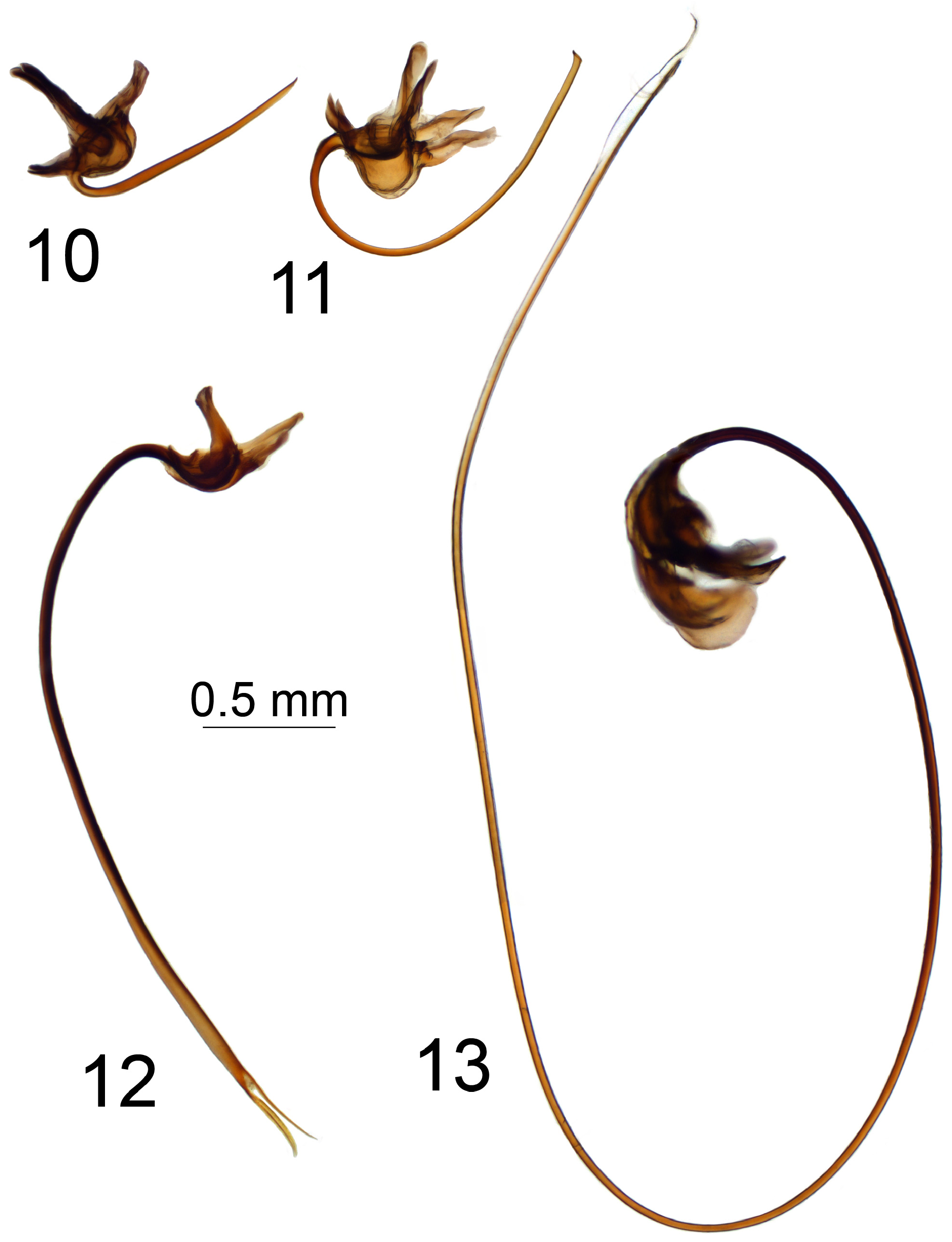
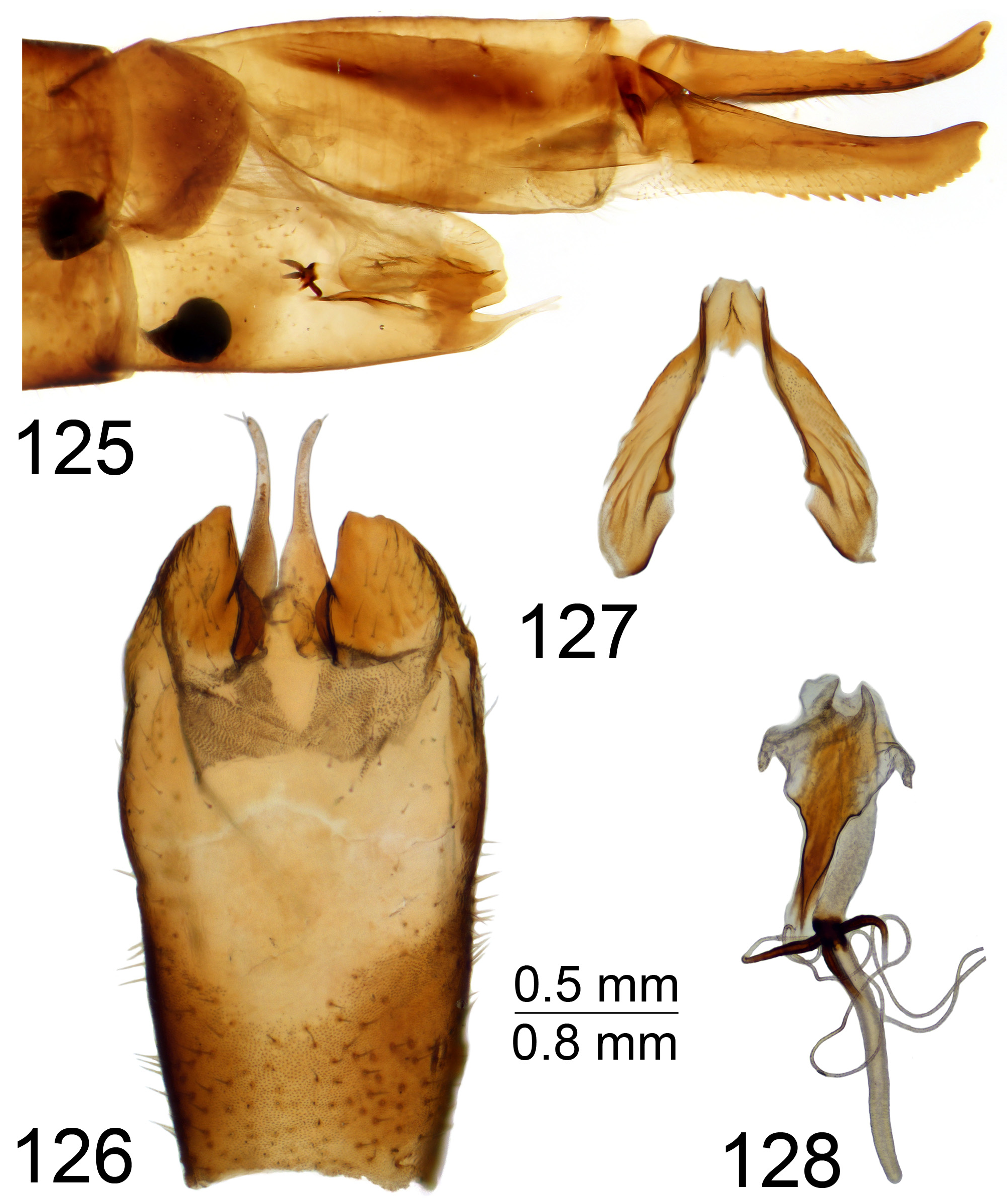
The gonocoxal fragment (sclerites sp1 and sp2, Neumann 1958; genital bridge, Dobrotworsky 1968) differs among T. ( Vestiplex ) species and is composed of lateral and medial sclerites ( Brodo 2018) with discussion of interpretation in Gelhaus (2005). The medial sclerite was designated sclerite sp1 by Neumann (1958) and as central part of genital bridge by Tangelder (1985). A pair of lateral sclerites were designated sclerites sp2 by Neumann (1958) and as lateral parts of genital bridge by Tangelder (1985). As indicated by Neumann (1958), sclerites sp2 are paired structures, while sclerite sp1 is a separate unpaired structure. This structure also was refered to as a single sclerite by Tangelder (1985). However the medial sclerite appears to be composed of two separate sclerites medially membranously fused. Mongolian members of the virgatula species group have the medial sclerite composed of two separate sclerites anteriorly membranously fused ( Fig. 4 View FIGURES 4–9 ). Other Mongolian T. ( Vestiplex ) species have the medial sclerites anteriorly fused with the base; e.g., excisa , scripta and nubeculosa species groups ( Figs 5, 6 View FIGURES 4–9 , 133 View FIGURES 129–136 ). The Mongolian members of the leucoprocta group have specific medial sclerites with the posterior (distal) parts forming an arched structure, the base developed into an anterior apodeme, with an additional posterior apodeme in T. (V.) balioptera , T. (V.) laccata and T. (V.) tchukchi ( Figs 8, 9 View FIGURES 4–9 ). The lateral sclerite in its simplest form is a very small and sometimes an unrecognizable structure attached to the dorsomedial edge of the gonocoxite. This state occurs in the virgatula , excisa and nubeculosa species groups. The Mongolian members of the scripta group have a small, slightly bilobed lateral sclerite ( Fig. 6 View FIGURES 4–9 ). The outer lobe is attached to the dorsomedial edge of the gonocoxite, with the inner lobe short and free in T. (V.) scripta or is attached to the mesal part of the gonocoxite membranously in T. (V.) pallitergata ( Fig. 7 View FIGURES 4–9 ). The members of the leucoprocta group have the lateral sclerite large and bilobed. Its outer lobe is attached to the dorsomedial edge of the gonocoxite, while the inner lobe is attached to the mesal part of the gonocoxite ( Figs 8, 9 View FIGURES 4–9 ). The sperm pump is usually flattened; compressor apodeme with or without median incision, and anterior immovable apodeme variable in shape from short to broadly flattened plate. The length of the aedeagus varies from short or relatively short ( Fig. 10 View FIGURES 10–13 ), moderately long ( Fig. 11 View FIGURES 10–13 ), to elongate or very long ( Figs 12, 13 View FIGURES 10–13 ). The distal part of the aedeagus is variously shaped, appearing trident ( Figs 170 View FIGURES 163–170 , 181 View FIGURES 174–181 ), funnel ( Figs 35 View FIGURES 28–35 , 136 View FIGURES 129–136 , 149 View FIGURES 149–151 ) or shovel-shaped ( Figs 46 View FIGURES 39–46 , 57 View FIGURES 50–57 , 70 View FIGURES 63–70 , 82 View FIGURES 75–82 , 124).
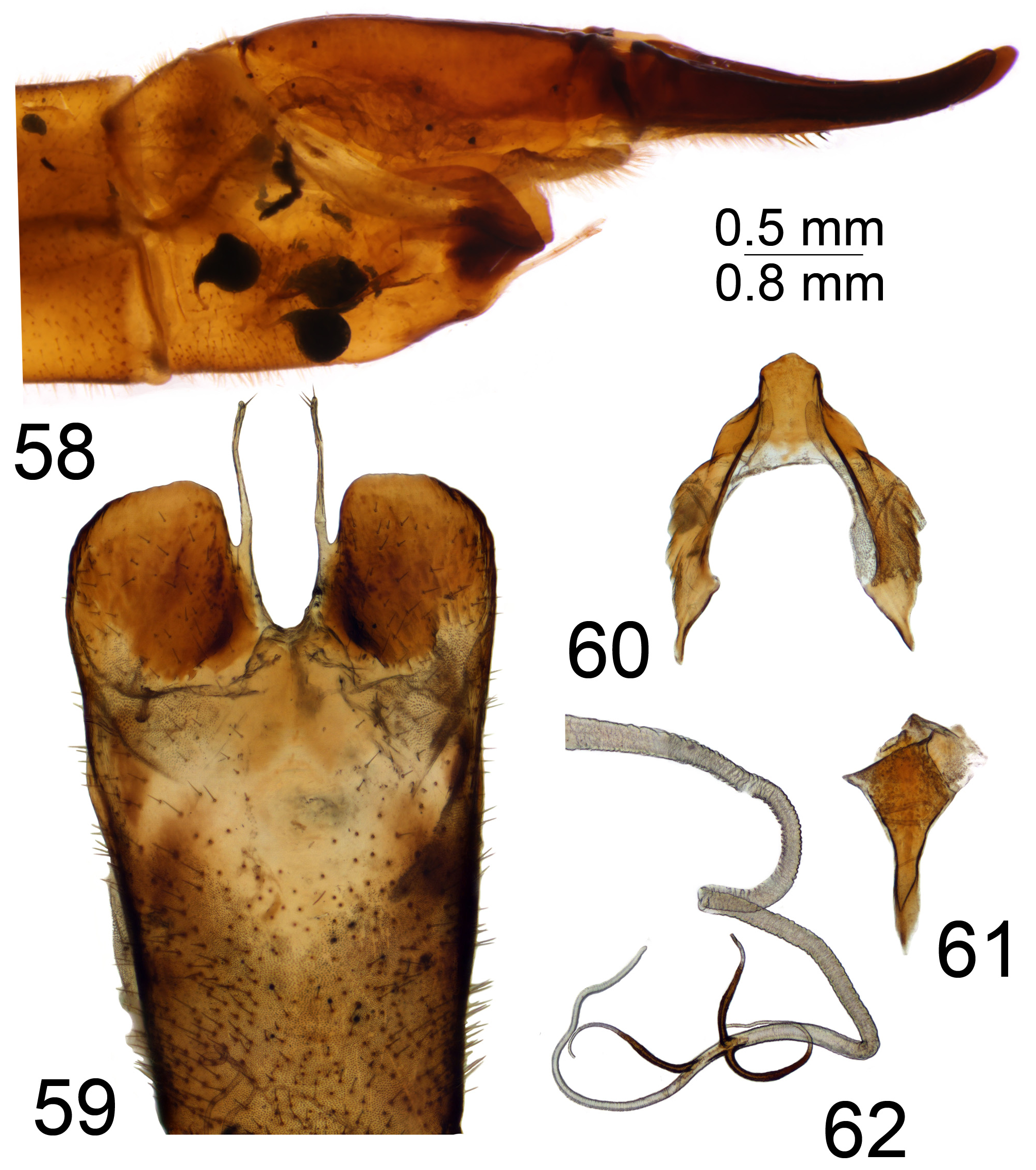
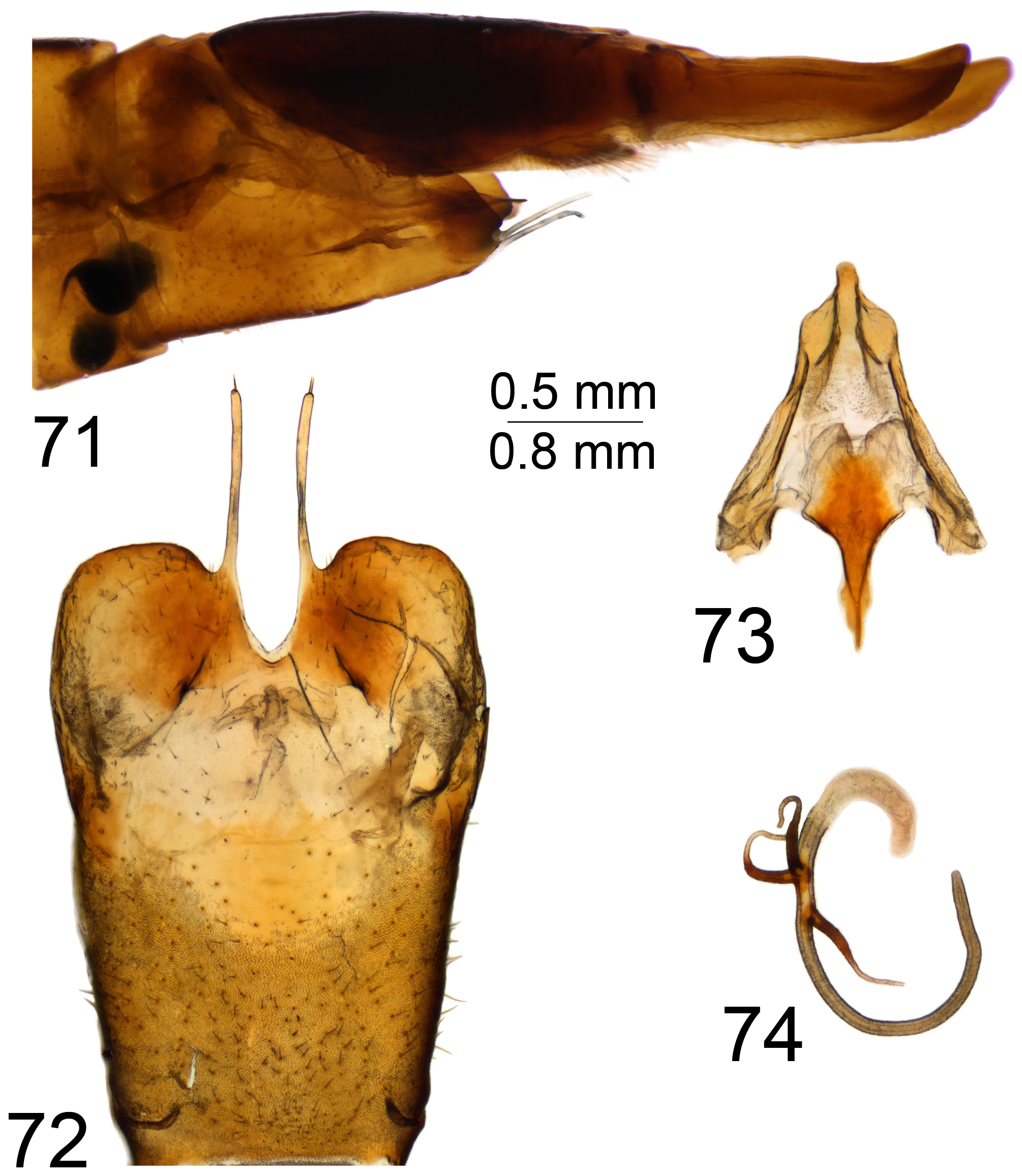
The female cercus is strong and powerfully constructed, heavily sclerotised, oriented horizontally. The outer (ventral) margin bears small to distinct, rough serration. The hypovalva of sternite 8 is with apical filaments ( Figs 37 View FIGURES 36–38 , 48 View FIGURES 47–49 , 59 View FIGURES 58–62 , 72 View FIGURES 71–74 , 84 View FIGURES 83–85 , 94 View FIGURES 93–95 , 104 View FIGURES 103–106 , 115 View FIGURES 114–116 , 126 View FIGURES 125–128 , 138 View FIGURES 137–141 , 150 View FIGURES 149–151 , 161 View FIGURES 160–162 ) or with short and stout projections in the virgatula species group ( Figs 172 View FIGURES 171–173 , 183 View FIGURES 182–184 ). Some female Mongolian T. ( Vestiplex ) could be distinguished by the remarkable bursa copulatrix and associated structures; e.g., members of leucoprocta species group have the basal part of the spermathecal ducts modified into sclerotised processes ( Figs 49 View FIGURES 47–49 , 62 View FIGURES 58–62 , 74 View FIGURES 71–74 , 95 View FIGURES 93–95 , 116 View FIGURES 114–116 ), with blackened wall on connection sites with bursa copulatrix ( Figs 85 View FIGURES 83–85 , 106 View FIGURES 103–106 , 128 View FIGURES 125–128 ). In other cases the spermathecal ducts are membranous ( Figs 38 View FIGURES 36–38 , 151 162 View FIGURES 149–151 View FIGURES 152–159 View FIGURES 160–162 , 173 View FIGURES 171–173 , 184 View FIGURES 182–184 ), while the wall of the bursa copulatrix including cul-de-sac is distinctly sclerotised; e.g., T. (V.) nubeculosa ( Fig. 141 View FIGURES 137–141 ).
Three spermathecae are present. The spermathecal shape varies and it is more or less spherical (e.g., T. (V.) longitudinalis , T. (V.) virgatula and T. (V.) pallitergata ( Figs 14–16 View FIGURES 14–27 , respectively)), lightly broadened at base (e.g., T. (V.) laccata , T. (V.) balioptera , T. (V.) nubeculosa ( Figs 17–19 View FIGURES 14–27 , respectively)) and with base distinctly broadened, oval or nearly pear-shaped (e.g., T. (V.) scripta , T. (V.) kiritshenkoi , T. (V.) mediovittata , T. (V.) subcentralis , T. (V.) sintenisi , T. (V.) coronifera , T. (V.) tchukchi and T. (V.) leucoprocta ( Figs 20–27 View FIGURES 14–27 , respectively)).





















No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Tipula
| Starkevich, Pavel, Podenas, Sigitas & Gelhaus, Jon K. 2020 |
Tipula
| Alexander, C. P. 1965: 355 |
| Savchenko, E. N. 1964: 132 |
| Mannheims, B. 1953: 116 |
| Alexander, C. P. 1935: 117 |
| Alexander, C. P. 1934: 396 |
| Edwards, F. W. 1931: 79 |
| Bezzi, M. 1924: 230 |