
Otobothrium curtum ( Linton, 1909 ) Dollfus, 1942
|
publication ID |
https://doi.org/ 10.5281/zenodo.178495 |
|
DOI |
https://doi.org/10.5281/zenodo.5693666 |
|
persistent identifier |
https://treatment.plazi.org/id/03E23E48-FFDC-664E-7EDF-FC9D9ED9FC77 |
|
treatment provided by |
Plazi |
|
scientific name |
Otobothrium curtum ( Linton, 1909 ) Dollfus, 1942 |
| status |
|
Otobothrium curtum ( Linton, 1909) Dollfus, 1942 View in CoL
( Figs. 9–20 View FIGURES 9 – 14 View FIGURES 15 – 20 , 51–52 View FIGURES 51 – 54 )
Syn. Rhynchobothrium curtum Linton, 1909
Types: USNPC 9000.
Type host: Galeocerdo cuvier (Péron & Lesueur, 1822) .
Type locality: Tortugas, Florida, USA, 2. vii. 1906, coll. E. Linton.
Site in host: spiral valve.
Material examined: From Galeocerdo cuvier (Péron & Lesueur, 1822) : types; 31 specimens, Baie de Prony, New Caledonia, from two sharks ( MNHN JNC 1207H1–H26, 1414A1–A20); 10 specimens, Mackay, Qld, Australia ( SAM 29088).
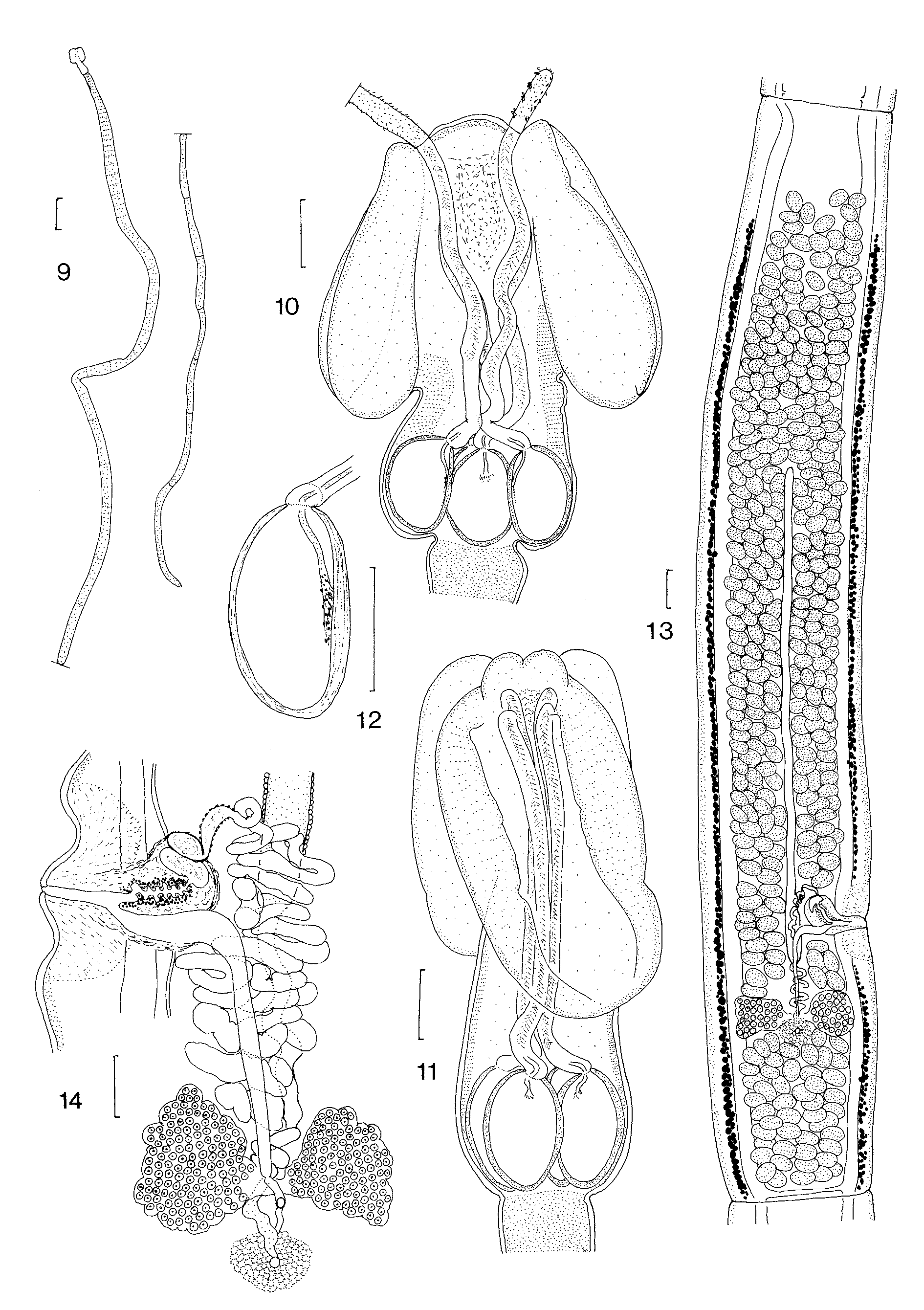
Re-description: Types: all immature, length 2.45–5.42 (3.47, n=5) mm, with 9–19 (13, n=5) segments; scolex length 260–340 (300, n=5); maximum width at level of bulbs 80–120 (100, n=5); pars bothrialis 220– 290 (240, n=5) long, bothrial width 120–190 (160, n=3); pars vaginalis 180–240 (210, n=5) long; bulbs 70–98 (82, n=5) long, 30–50 (45, n=5) wide. Most mature segment 1.52 mm long, 350 wide; genital pore to posterior end 380; genitalia poorly developed; testes 25 post-ovarian, 1 testis between ovary and genital atrium; hundreds of testes (not countable) anterior to ovaries.
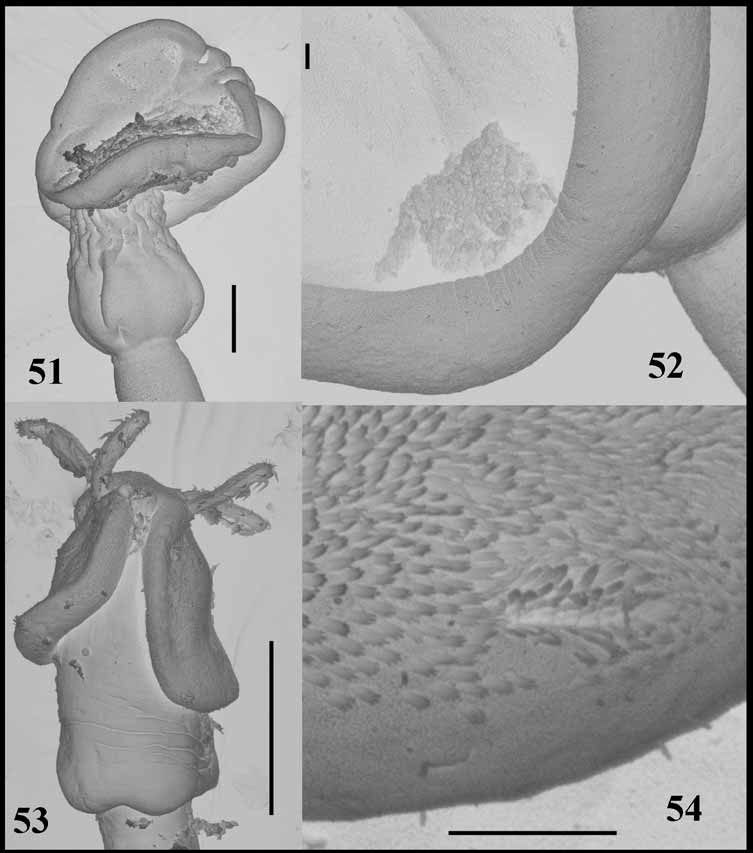
Specimens from New Caledonia: Longest cestode 37 mm, maximum width 500, with 65 segments; hyperapolytic. Scolex acraspedote, 500–700 (585, n=10) long; maximum width in pars bulbosa 190–260 (230, n=10). Two narrow, elongate bothria extending to anterior extremity of bulbs; pars bothrialis 310–490 (395, n=10), width of bothria 250; bothrial pits absent. Pars vaginalis equal in length to pars bothrialis, 310–510 (395, n=10) long; tentacle sheaths straight in anterior part, sinuous anterior to bulbs. Bulbs ovoid, 140–160 (154, n=10) long, 80–100 (89, n=10) wide; prebulbar organ absent; retractor muscle originates in anterior part of bulb, insertion continued posteriorly by band of nucleated tissue; pars post-bulbosa absent.
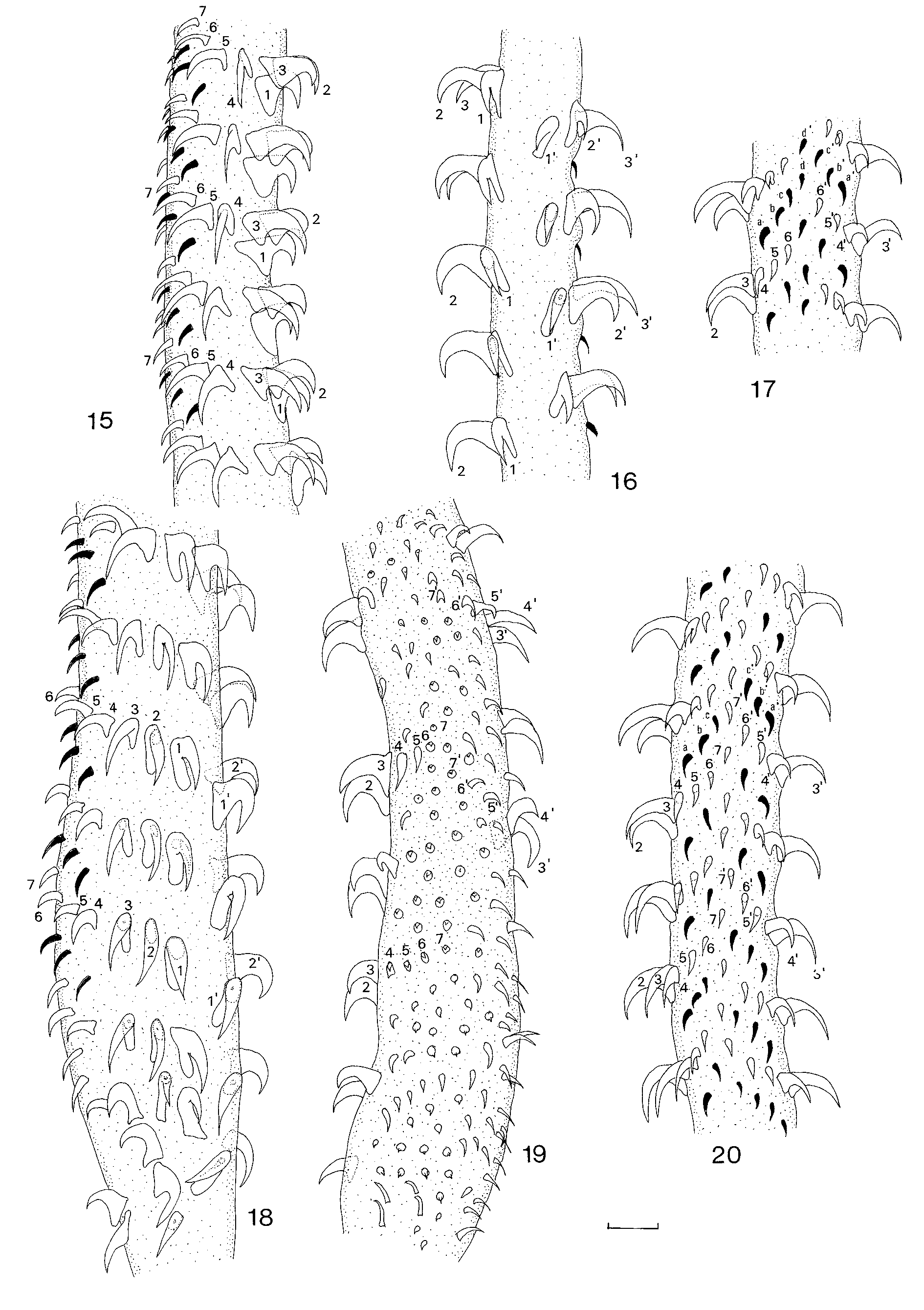
Everted tentacles up to 670 long; slight basal swelling present; diameter at base 25–38 (30, n=10), diameter in distal region 15–25 (22, n=10). Armature heteroacanthous, atypical, heteromorphous; hooks hollow. Hook files begin on internal surface of tentacle, terminate on external surface; 7 hooks per row; space present between hook files 1 and 1’ on internal surface of tentacle. Distinctive basal armature absent. Hooks 1(1’) large, uncinate, with broad base, 10–12 (11, n=5) long, base 8–10 (9, n=5); hooks 2(2’), erect, with broad blade and relatively long base, 10–14 (12, n=5) long, base 7–8 (7.5, n=5); hooks 3(3’) longer, slender, falcate, with shorter base, 13–16 (15, n=5) long, base 6–8 (7, n=5); hooks 4(4’) shorter, falcate, with smaller bases, 8– 11(10, n=5) long, base 4–7 (5, n=5); hooks 5(5’) short, spiniform, 5–13 (9, n=5) long, base 3–5 (4, n=5); hooks 6(6’) short, spiniform, 6–8 (7, n=5) long, base 1–3 (2, n=5); hooks 7(7’) short, spiniform, 5–6 (6, n=5) long, base 1–2 (1.6, n=5). In metabasal region, single intercalary row of hooks between each principal row; intercalary hooks spiniform, 3–8 (6, n=5) long, base 1–2 (1.4, n=5); in basal region on external surface, 2– 3 intercalary rows with 2– 3 intercalary hooks per row.
Mature segments elongate, acraspedote, 1.91–2.87 (2.23, n=5) mm long, maximum width 280–550 (450, n=5); segment length: width ratio 4.1–7.0 (5.2, n=5); genital pores alternate irregularly, 450–660 (530, n=5) from posterior end of segment. Terminal genital ducts visible only in hyperapolytic segments. Deeply staining areas anterior and posterior to genital atrium. Hermaphroditic sac thin-walled, 220–300 (260, n=5) long, 110– 180 (146, n=5) wide. Cirrus joins vagina in distal part of hermaphroditic sac; cirrus corrugated, muscular, unarmed, leads to small internal seminal vesicle at proximal pole of hermaphroditic sac; vesicle not visible in all specimens, 30–70 (57, n=3) long, 20–40 (28, n=3) wide; external seminal vesicle absent, but initial coils of vas deferens with distinctive cellular wall. Vas deferens coiling posteriorly to ovarian isthmus. Testes intervascular, arranged in single layer throughout medulla, not reaching anterior part of segment; testes 45–65 (56, n=10) long, 33–58 (42, n=10) wide; total number of testes per segment 289–345 (316, n=5), with 49–57 (53, n=5) post-ovarian, 3–6 (5, n=5) post-vaginal, and 233–291 (259, n=5) prevaginal/aporal. Vagina penetrates proximal pole of hermaphroditic duct, then runs posteriorly to ovarian isthmus; seminal receptacle absent. Ovary bilobed in dorso-ventral view, tetra-lobed in transverse section, lobes 330–550 (390, n=5) long, 160– 230 (210, n=5) wide; Mehlis’ gland posterior to ovarian isthmus, 150-220 (170, n=5) in diameter. Vitelline follicles circum-medullary, 15–30 (20, n=10) in diameter. Uterine duct coils anteriorly from Mehlis’ gland to level of hermaphroditic sac; uterus extends from this point to anterior quarter of segment, not reaching anterior extent of testes; uterine pore absent. Ventral osmoregulatory canal 13–43 (23, n=5) in diameter, with transverse canal 13–38 (19, n=5) connecting ventral canals at posterior end of each segment; dorsal canal tiny, c. 3 in diameter, internal to ventral canal.
Hyperapolytic segments 3.8–8.4 (5.8, n=5) mm long, 640–840 (790, n=5) wide; length to width ratio 5.1– 10.3 (7.3, n=5); genital pore 680–1780 (1170, n=5) from posterior end. Single pre-gravid segment 5.02 mm long, maximum width 250; ovarian lobes 800 long, 200 wide, Mehlis’ gland 180 in diameter; uterus elongate, saccate, without lateral lobes; eggs yellow, ovoid, 45–53 (49, n=10) long, 28–30 (29, n=10) wide.
Remarks. Linton (1909) gave a very brief description of this species based on specimens collected from a tiger shark, Galeocerdo cuvier , off the eastern coast of the United States at Dry Tortugas, Florida. The specimens he described were immature (10 mm long in his description, maximum length 5.42 in type specimens) and no tentacles were everted. He also reported plerocerci from the serranids Epinephelus striatus (Bloch, 1792) and Mycteroperca bonaci (Poey, 1860) . Linton (1909) allocated the species to Rhynchobothrium rather than to his genus Otobothrium because it did not possess observable bothrial pits. Dollfus (1942) included the species in Otobothrium , but doubted its validity, and suggested that it may be a synonym of O. crenacolle . He stated, correctly, that it was not uncommon to encounter difficulties in finding the bothrial pits in the known species of Otobothrium . He considered the other characters used by Linton (1909) to distinguish this species, the lack of divergence of the bulbs and the supposed "lateral" bothria to be due to intraspecific variation in the former case, or a simple error in the latter case. Yamaguti (1959) and Schmidt (1986) listed it as a species of Otobothrium , however, Palm (2004) synonymised the species with O. crenacolle with no explanation for his action.
The new material from Galeocerdo cuvier from Australia and New Caledonia demonstrates that O. curtum is indeed a valid species and is apparently specific to tiger sharks. It is readily distinguished from congeners in its narrow, elongate bothria which are, on average, as long as the pars vaginalis, in the size and shape of its mature segments and in the number of testes per segment (289–345 compared with 98–139 in O. crenacolle ). Differences exist also in the armature of the metabasal region of the tentacle where there are seven hooks per principal row compared with smaller numbers in most related species. However, depending on how the hooks are numbered, there may be seven hooks in each principal row with an intercalary row of three hooks ( Fig. 20 View FIGURES 15 – 20 ). In this case, hooks 7 and 7' form a file running along the middle of the external surface of the tentacle. An alternative method of numbering the hooks ( Fig.17 View FIGURES 15 – 20 ) would allow six hooks in each principal row, with four hooks in each intercalary row, the fourth hooks d and d' forming a file on the external surface of the tentacle. Since there are no morphological differences between the terminal hooks of the principal rows and the intercalary hooks, either method of numbering the hooks is considered to be equally appropriate.
As indicated originally by Linton (1909), there are no bothrial pits. In this study, they were not detected either by light or scanning electron microscopy. Dollfus' (1942) general observation that bothrial pits may be difficult to detect is undisputed, but the wide range of specimens available for this study and the use of scanning electron microscopy confirms that, in this species, bothrial pits and/or modified microtriches do not exist. In the related genus Symbothriorhynchus Yamaguti, 1952 , Palm (2004) demonstrated that the bothrial pits were very small, only readily detectable using electron microscopy and were identifiable based on the presence of modified microtriches. However, in the case of O. curtum , even scanning electron microscopy failed to detect the existence of bothrial pits or modified microtriches. Although this is a key distinguishing feature of the genus, O. curtum is retained provisionally within the genus.
The mature and gravid segments described in this species were clearly attributable to O. curtum since the entire contents of the spiral valves of two tiger sharks from New Caledonia were examined microscopically and the only trypanorhynch cestode present was O. curtum .
Australia and New Caledonia are new locality records for this species.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.